


营养盐是海洋微藻生长的重要生态因子,影响着藻类生物的生理活动和生化组成,以及其种群的生长和繁殖[27-28]。其中,氮、磷营养盐是海水的主要组分,近年来,有关氮、磷限制对海洋微藻生长及生化组成的影响已有许多进展。研究表明,氮是藻细胞中光合作用的必需元素,当氮营养盐不足时,藻细胞内叶绿素总量减少,光反应中心蛋白合成减少,光能的利用效率降低,影响微藻的生长与繁殖[29];无机磷在微藻细胞中合成有机磷化合物,参与合成ATP、GTP、辅酶等光合作用所需的底物或调节物[30-31]。夏建荣等[29]研究发现,小新月菱形藻细胞的比生长速率和稳定期细胞密度随氮浓度的增加而增加,说明无机氮浓度的升高有利于细胞分裂;同时,磷浓度越低,小新月菱形藻的比生长速率越低。梁晶晶等[32]研究了氮、磷、铁对三角褐指藻诱变株MP-2生长、总脂及脂肪酸的影响,结果表明该诱变株生长最适氮浓度为20 mg·L -1,当氮浓度为30 mg·L -1时,总脂积累量最高;生长最适磷浓度为2.5 mg·L -1,当磷浓度为1.0 mg·L -1时总脂积累量最高。然而,对于不同氮、磷营养盐条件下岩藻黄素的积累研究仍较少。本研究拟在f/2培养基的基础上,研究不同的氮、磷营养盐对威氏海链藻生长和岩藻黄素积累的影响,通过HPLC定量检测岩藻黄素含量,探讨氮、磷营养盐对威氏海链藻生长的影响,以优化培养条件,提高岩藻黄素产量。
1 材料与方法
1.1 材料
1.1.1 实验材料
实验所需威氏海链藻(Thalassiosira weissflogii)购于上海光语生物科技有限公司;采用f/2培养基培养,待用。
1.1.2 实验试剂
无水乙醇、甲醇、丙酮等试剂为国药分析纯;色谱级乙腈、色谱级甲醇购于德国默克公司;岩藻黄素标准品为液相色谱级。
1.1.3 主要仪器设备
真空冷冻干燥机(松原华兴LGJ-12)、超声波清洗器(昆山禾创KQ5200E)、分析天平(奥豪斯CP114)、高速离心机(艾本德5424R)、台式高速离心机(赛默飞世尔)、涡旋仪(塞维尔生物MV-100)、自动细胞计数仪(上海睿钰生物有限公司)、高效液相色谱(美国Waters e2695)等。
1.2 方法
1.2.1 氮营养盐条件设计
配制f/2缺氮培养基,选用硝酸钠、硫酸铵、尿素作为氮营养盐,根据f/2培养基配方及参考梁晶晶等[32]对三角褐指藻的生长研究,设置氮的浓度为0、0.882、1.323、1.764、2.205 mmol·L -1,每种处理设3个平行,以正常f/2培养基培养的藻作为对照。培养条件:23℃,75 μmol·m-2·s -1,全天光照,每天手摇2~3次。
1.2.2 磷营养盐条件设计
上述优化的氮营养盐配制f/2缺磷培养基,选用磷酸二氢钠和磷酸二氢钾作为磷营养盐,磷的浓度设置为0.018、0.036、0.054、0.072 mmol·L -1,每种处理设3个平行,以正常f/2培养基培养的藻作为对照。培养条件:23℃,75 μmol·m-2 ·s -1,全天光照,每天手摇2~3次。
1.2.3 威氏海链藻细胞密度曲线的建立
接种后,每日于超净台定时取样,卢戈氏碘液固定后,通过自动细胞计数仪计数,并绘制细胞密度曲线。
1.2.4 威氏海链藻生物量测定
待生长至稳定后期,藻液8 000 r/min离心5 min,蒸馏水洗涤2~3次以除去海水中的盐离子,于-20℃冰箱中放置3 h后转移到-80℃冰箱中预冷。从-80℃冰箱中取出,置于真空冷冻干燥机中,冷冻干燥40 h。结束后用分析天平称重,计算可得威氏海链藻生物量(dw)=干燥后总重量(m2)-离心管重量(m1)。
1.2.5 威氏海链藻岩藻黄素的提取与检测
在无光/弱光环境中,加入100 mL/g干重4℃预冷的混合溶液(丙酮∶甲醇=1∶1),涡旋震荡20~30 s混匀后,置于4℃冰箱避光浸提45 min。取出后10 000 r/min离心5 min,此时离心管中的上清液即为岩藻黄素粗提液。通过0.22 μm有机微孔滤膜过滤,装入2 mL棕色样品瓶中。使用高效液相色谱仪Waters e2695对样品进行检测,得到每个样品中岩藻黄素的含量(μg·mL-1)。
1.2.6 岩藻黄素标准曲线的建立
以色谱级甲醇为溶剂,溶解岩藻黄素标准品,分别配置浓度为1、5、10、20、40、60、80、100 μg·mL-1的岩藻黄素标准液,HPLC检测后根据浓度和峰面积建立标准曲线。
2 结果与分析
2.1 岩藻黄素标准曲线
图1
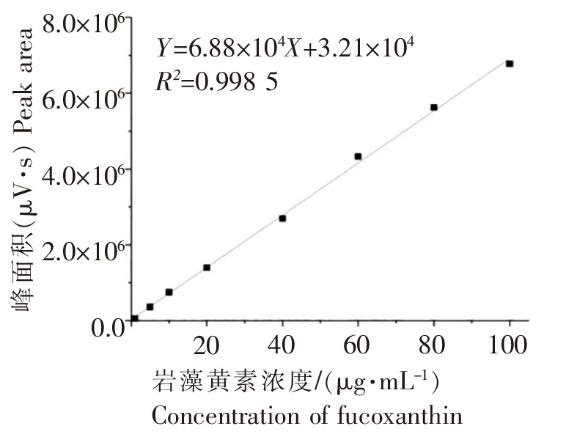
2.2 氮营养盐对威氏海链藻生长和岩藻黄素含量的影响
2.2.1 不同浓度硝酸钠对威氏海链藻生长和岩藻黄素含量的影响
不同浓度的硝酸钠对威氏海链藻生长和岩藻黄素含量的影响如图2所示。添加了硝酸钠的4组威氏海链藻在接种后的第6天,细胞密度基本上都达到最高值;而缺氮组在第4天就达到最高值,且远低于其他4组(图2A)。当硝酸钠浓度为1.764 mmol·L -1时,生物量最高,但与其他3组添加了硝酸钠的相比,无显著性差异(P>0.05)(图2B)。当硝酸钠浓度分别为0.882、1.764 mmol·L -1时,每g干重威氏海链藻中岩藻黄素的含量较高,两组之间无显著性差异(P>0.05),但含量最高组(硝酸钠浓度1.764 mmol·L -1)与其他2组(硝酸钠浓度分别为1.323、2.205 mmol·L -1)相比,存在显著性差异(P<0.05)(图2C)。当硝酸钠浓度为1.764 mmol·L -1时,每L威氏海链藻藻液中岩藻黄素的总量较高,与其他4组相比,均存在显著性差异(P<0.05)(图2D)。在不添加硝酸钠时,威氏海链藻生物量、岩藻黄素含量及总量均与其他4组存在显著性差异,威氏海链藻的生长和岩藻黄素积累受到明显影响(图2)。
图2
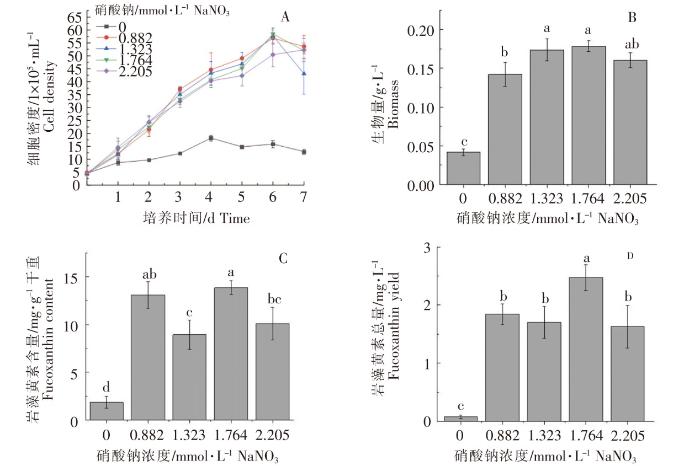
图2
不同浓度硝酸钠对威氏海链藻细胞密度(A)、生物量(B),岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.2
The effects of sodium nitrate in different concentrations on cell density (A), biomass (B), fucoxanthin content (C) and fucoxanthin yield (D) of Thalassiosira weissflogii
2.2.2 不同浓度硫酸铵对威氏海链藻生长和岩藻黄素含量的影响
以正常f/2培养基(硝酸钠浓度0.882 mmol·L-1)为对照,不同浓度硫酸铵对威氏海链藻的生长和岩藻黄素含量的影响如图3所示。不同硫酸铵浓度培养的威氏海链藻基本上均在接种后的第6天,细胞密度达到最高值;当硫酸铵浓度为0.441 mmol·L -1时,威氏海链藻细胞密度较大,且高于对照组,较低浓度的硫酸铵有助于威氏海链藻细胞密度的增长(图3A)。当硫酸铵浓度为0.441 mmol·L -1时,生物量最高,与较高浓度硫酸铵处理(硫酸铵浓度分别为0.882、1.103 mmol·L -1)及对照组相比,存在显著性差异(P<0.05)(图3B)。当硫酸铵浓度分别为0.441、0.662 mmol·L -1时,每g干重威氏海链藻中岩藻黄素的含量较高,两组之间无显著性差异(P>0.05),但与最高浓度处理组(硫酸铵浓度为1.103 mmol·L -1)相比,存在显著性差异(P<0.05)(图3C)。当硫酸铵浓度为0.441 mmol·L -1时,每L威氏海链藻藻液中岩藻黄素的总量较高,与高浓度处理组(硫酸铵浓度为0.882、1.103 mmol·L -1)相比,存在显著性差异(P<0.05)(图3D)。由图3可以看出,与对照组相比,以硫酸铵作为氮营养盐培养威氏海链藻,低浓度的硫酸铵有利于威氏海链藻生长及岩藻黄素积累,而在不添加硫酸铵时,威氏海链藻的生长和岩藻黄素积累则受到明显影响。
图3
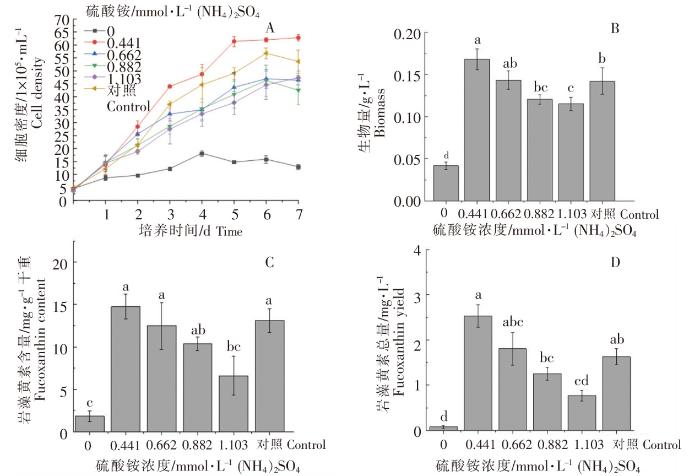
图3
不同浓度硫酸铵对威氏海链藻细胞密度(A)、生物量(B),岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.3
The effects of ammonium sulfate in different concentrations on cell density (A), biomass(B), fucoxanthin content (C) and fucoxanthin yield (D) of Thalassiosira weissflogii
2.2.3 不同浓度尿素对威氏海链藻生长和岩藻黄素含量的影响
以正常f/2培养基(硝酸钠浓度0.882 mmol·L-1)为对照,不同浓度尿素对威氏海链藻的生长和岩藻黄素含量的影响如图4所示。不同尿素浓度培养的威氏海链藻基本上均在接种后的第6天,细胞密度达到最高值;不同浓度尿素对威氏海链藻细胞密度的影响不明显。当尿素浓度为0.882 mmol·L -1时,细胞密度较大(图4A);生物量最高,与尿素浓度为0.662、1.103 mmol·L -1的实验组及对照组相比,存在显著性差异(P<0.05)(图4B);每g干重威氏海链藻中岩藻黄素含量较高,与其他组相比,均存在显著性差异(P<0.05)(图4C);每L威氏海链藻藻液中岩藻黄素总量较高,与其他组相比,均存在显著性差异(P<0.05)(图4D)。由图4可以看出,与对照组相比,以尿素作为氮营养盐培养威氏海链藻,浓度为0.882 mmol·L -1的尿素明显有利于威氏海链藻生长及岩藻黄素积累,在不添加氮营养盐时,威氏海链藻的生长和岩藻黄素积累受到明显影响。
图4
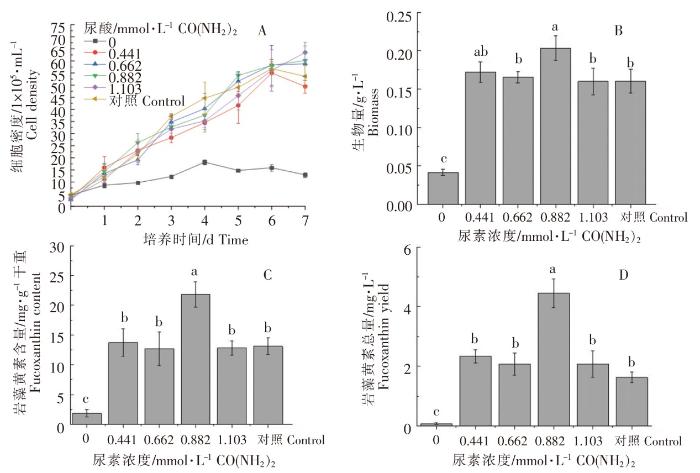
图4
不同浓度尿素对威氏海链藻细胞密度(A)、生物量(B)、岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.4
The effects of urea in different concentrations on cell density (A), biomass(B), fucoxanthin content(C) and fucoxanthin yield (D) of Thalassiosira weissflogii
2.2.4 三种最佳浓度的氮营养盐对威氏海链藻生长和岩藻黄素含量影响的比较
根据前述比较,选取三种最佳浓度的氮营养盐进行数据分析,结果如图5所示。不同最佳浓度的氮营养盐培养的威氏海链藻基本上均在接种后的第6天,细胞密度达到最高值;其中在硫酸铵浓度为0.441 mmol·L -1时,威氏海链藻细胞密度最高(图5A)。在0.882 mmol·L -1尿素处理下,生物量最高,与对照组相比存在显著性差异(P<0.05)(图5B);每g干重威氏海链藻中岩藻黄素含量最高,与其他2组及对照组相比,均存在显著性差异(P<0.05)(图5C);每L威氏海链藻藻液中岩藻黄素总量最高,与其他2组及对照组相比,均存在显著性差异(P<0.05)(图5D)。由图5可以看出,以0.882 mmol·L -1尿素为氮营养盐,有利于威氏海链藻的生长及岩藻黄素积累。在0.441 mmol·L -1硫酸铵处理下,细胞密度最大,但在0.882 mmol·L -1尿素处理下,生物量最大,结合岩藻黄素含量结果,分析可能的原因为在0.441 mmol·L -1硫酸铵处理下的威氏海链藻生长速度较快,但油脂等代谢产物较少,细胞个体较小,导致生物量较低。
图5
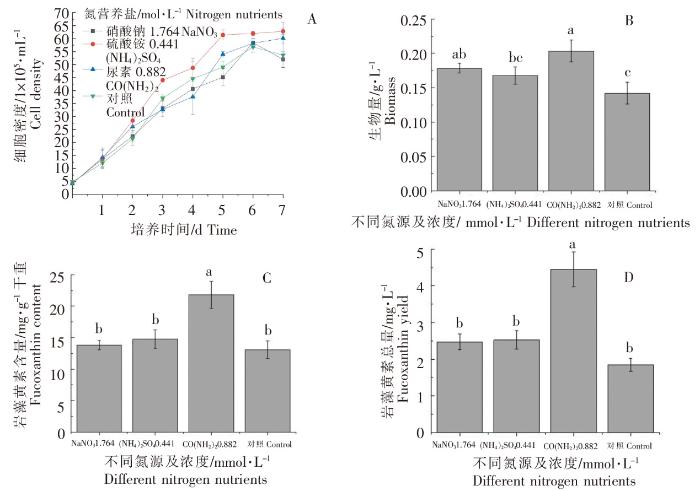
图5
不同处理对威氏海链藻细胞密度(A)、生物量(B),岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.5
The effects of different treatment on cell density (A), biomass (B), fucoxanthin content(C) and fucoxanthin yield (D) of Thalassiosira weissflogii
2.3 磷营养盐对威氏海链藻生长和岩藻黄素含量的影响
2.3.1 不同浓度磷酸二氢钠对威氏海链藻生长和岩藻黄素含量的影响
图6
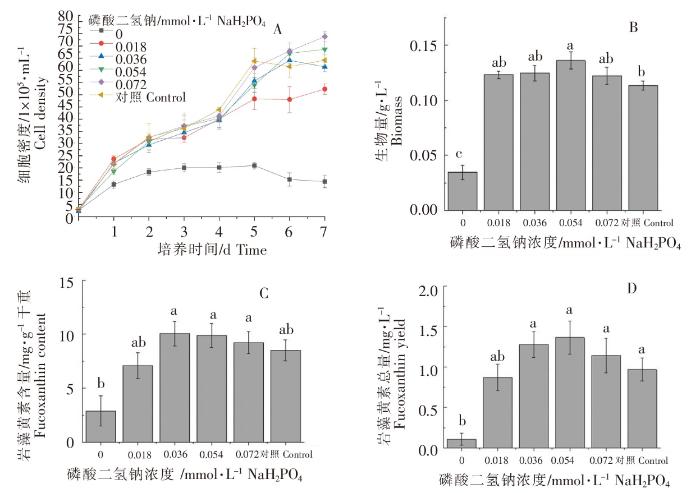
图6
不同浓度磷酸二氢钠对威氏海链藻细胞密度(A)、生物量(B),岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.6
The effects of sodium dihydrogen phosphate in different concentrations on cell density (A), biomass(B) fucoxanthin content (C) and fucoxanthin yield (D) of Thalassiosira weissflogii
2.3.2 不同浓度磷酸二氢钾对威氏海链藻生长和岩藻黄素含量的影响
不同浓度磷酸二氢钾对威氏海链藻的生长和岩藻黄素含量的影响如图7所示。添加了磷酸二氢钾的4组和对照组在接种后的第5、6天,威氏海链藻的细胞密度达到最高值,较低浓度的磷酸二氢钾(浓度为0.018 mmol·L -1)不利于威氏海链藻细胞密度的增长(图7A)。不同浓度磷酸二氢钾对生物量的影响不明显,不存在显著性差异(P>0.05)(图7B)。当磷酸二氢钾浓度为0.054 mmol·L -1时,每g干重威氏海链藻中岩藻黄素含量较高,与其他组相比,均存在显著性差异(P<0.05)(图7C)。当磷酸二氢钾浓度为0.054 mmol·L -1时,每L威氏海链藻藻液中岩藻黄素总量较高,与其他组相比,均存在显著性差异(P<0.05)(图7D)。由图7可以看出,0.054 mmol·L -1磷酸二氢钾有利于岩藻黄素的积累。
图7
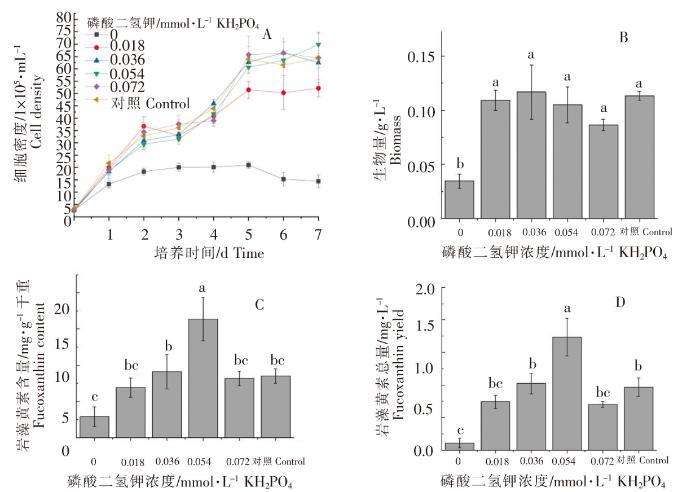
图7
不同浓度磷酸二氢钾对威氏海链藻细胞密度(A)、生物量(B),岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.7
The effects of potassium dihydrogen phosphate in different concentrations on cell density (A),biomass (B). fucoxanthin content (C) and fucoxanthin yield (D) of Thalassiosira weissflogii
2.3.3 两种最佳浓度磷营养盐对威氏海链藻生长和岩藻黄素含量影响的比较
根据前述比较结果,选取两种最佳浓度的磷营养盐进行数据分析,结果如图8所示。不同最佳浓度的磷营养盐培养的威氏海链藻基本上均在接种后的第6、7天,细胞密度达到最高值(图8A)。在0.054 mmol·L -1磷酸二氢钠处理下,生物量最高,与磷酸二氢钾组相比,存在显著性差异(P<0.05)(图8B)。在0.054 mmol·L -1磷酸二氢钾处理下,每g干重威氏海链藻中岩藻黄素含量最高,与其他组相比,均存在显著性差异(P<0.05)(图8C);每L威氏海链藻藻液中岩藻黄素总量最高,与其他组相比,均存在显著性差异(P<0.05)(图8D)。由图8可以看出,从细胞密度和生物量来看,以0.054 mmol·L-1磷酸二氢钠为磷营养盐,有利于威氏海链藻的生长,但0.054 mmol·L-1磷酸二氢钾更有利于岩藻黄素的积累。
图8
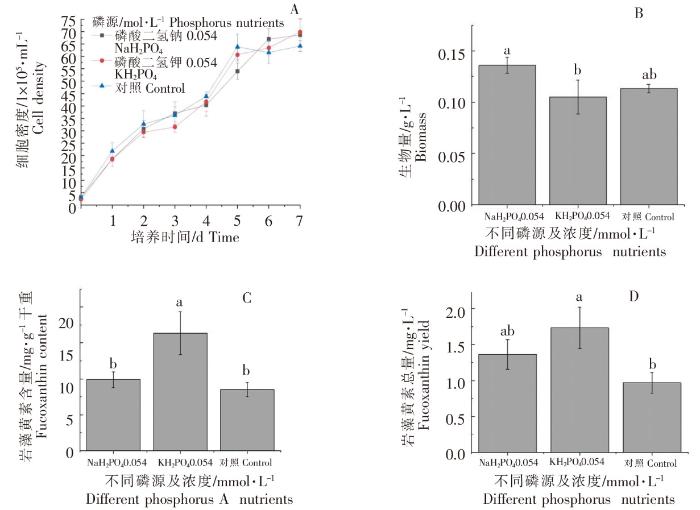
图8
不同处理对威氏海链藻细胞密度(A)、生物量(B),岩藻黄素含量(C)和岩藻黄素总量(D)的影响
Fig.8
The effects of different treatment on cell density (A), biomass(B), fucoxanthin content(C) and fucoxanthin yield (D) of Thalassiosira weissflogii
3 讨论与结论
本研究主要通过改变培养基中氮、磷营养盐浓度,比较其对威氏海链藻生长及岩藻黄素积累的影响。氮是威氏海链藻生长发育的重要生态因子,藻细胞能够对不同形态的氮营养盐进行选择性吸收和利用。徐兴莲等[33]研究发现,氮限制抑制了三角褐指藻的生长与光合活性,其叶绿素 a含量降低,PSⅡ光反应中心部分关闭。不同藻类对氮营养盐的利用存在差异,张青田等[34]研究了相同浓度下氯化铵和尿素对几种藻类生长的影响,发现氯化铵作为氮营养盐更有利于赤潮异弯藻、纤细角毛藻和亚历山大藻的生长,而尿素更有利于隐藻的生长。朱明等[35]研究表明,与硝酸盐相比,氨盐作为氮营养盐更有利于威氏海链藻的生长代谢,这是因为微藻对于氨盐的吸收引起培养基pH值的下降,对因光合作用导致的培养液pH值上升起到缓冲作用,而有利于微藻生长。本研究中,在相同氮浓度(0.441 mmol·L -1硫酸铵和 0.882mmol·L -1硝酸钠)培养下,硫酸铵更有利于威氏海链藻的生长与岩藻黄素的积累。窦勇等[5]研究发现,与无机氮营养盐相比,尿素更有利于提高威氏海链藻细胞密度和叶绿素 a 质量浓度,本研究中,0.882 mmol·L -1的尿素最适合威氏海链藻的生长及岩藻黄素的积累。另外,也有许多学者研究了氮磷比对威氏海链藻生长的影响,在相同氮磷比条件下,提高氮磷营养盐浓度有利于威氏海链藻的生长与光合作用[5];同时,刘皓等[36]研究发现,相比于磷营养盐,氮营养盐浓度对威氏海链藻的生长影响较大。本研究中,与磷营养盐相比,氮营养盐浓度对威氏海链藻生长及岩藻黄素积累影响较大,生物量增长与岩藻黄素积累量增加均更为明显,与前述研究结果基本一致。
本研究选取了硝酸钠、硫酸铵与尿素为氮营养盐,磷酸二氢钠与磷酸二氢钾为磷营养盐,分别设置了4个浓度梯度,实验结果表明:1)0.882 mmol·L -1尿素明显有利于威氏海链藻的生长及岩藻黄素的积累,与其他氮营养盐相比,存在显著性差异(P<0.05);生物量、岩藻黄素含量和岩藻黄素总量最高分别可达到 0.203 g·L-1、21.806 mg·g-1和4.449 mg·L-1,与对照组相比,生物量提高了45.4%,岩藻黄素含量提高了66.5%,岩藻黄素总量提高了173.1%。2)以上述优化的氮营养盐为基础,比较磷酸二氢钠和磷酸二氢钾两种磷盐对威氏海链藻生长及岩藻黄素积累的影响,发现0.054 mmol·L-1磷酸二氢钾在威氏海链藻生物量的积累上无显著性差异(P>0.05),但在岩藻黄素含量积累方面,与其他磷营养盐相比,存在显著差异(P<0.05)。与对照组相比,岩藻黄素含量提高了78.9%,岩藻黄素总量提高了92.1%。在缺氮、缺磷的条件下,威氏海链藻的生长及岩藻黄素的积累均受到严重影响。
本次实验研究了氮、磷营养盐对威氏海链藻生长的影响,在f/2培养基的基础上设置了不同的氮、磷营养盐及不同的浓度,为培养基优化及提高威氏海链藻岩藻黄素产量提供了理论基础。
参考文献
Carbon:Pheopigment ratios of zooplankton fecal pellets as an index of herbivorous feeding
[J].
Ingestion,growth and development of Penaeus indicus larvae as a function of Thalassiosira weissflogii cell concentration
[J].
The selection of an ideal diet for Ostrea edulis (L.)broodstock conditioning(part B)
[J].
Pigment signatures of some diatoms isolated from China seas
[J].
Extracts from New Zealand Undaria pinnatifida containing fucoxanthin as protenial fuctional biomaterials agains cancer in vitro
[J].
Proton-donative antioxidant activity of fucoxanthin with 1,1-Diphenyl-2-Picrylhydrazyl(DPPH)
[J].
Reactions between a carotenoid, fucoxanthin and 1,1-diphenyl-2-picrylhydrazyl were investigated both under anoxic and aerobic conditions. Fucoxanthin equimolarly reacted with 1,1-diphenyl-2-picrylhydrazyl under anoxic conditions. Under aerobic conditions, only a part of fucoxanthin consumed 1,1-diphenyl-2-picrylhydrazyl and the degree of reaction fluctuated with repeated trials. beta-Carotene or other carotenoids, beta-cryptoxanthin, zeaxanthin, licopen and lutein, were also examined in the reaction with 1,1-diphenyl-2-picrylhydrazyl under anoxic conditions. All these compounds scarcely reacted with 1,1-diphenyl-2-picrylhydrazyl.
Anti-obese property of fucoxanthin is partly mediated by altering lipid-regulating enzymes and uncoupling proteins of visceral adipose tissue in mice
[J].
Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice
[J].Fucoxanthin is a marine carotenoid found in edible brown seaweeds. We previously reported that dietary fucoxanthin attenuates the weight gain of white adipose tissue (WAT) of diabetic/obese KK- A(y) mice. In this study, to evaluate the antiobesity and antidiabetic effects of fucoxanthin and fish oil, we investigated the effect on the WAT weight, blood glucose, and insulin levels of KK- A(y) mice. Furthermore, the expression level of uncoupling protein 1 (UCP1) and adipokine mRNA in WAT were measured. After 4 weeks of feeding, 0.2% fucoxanthin in the diet markedly attenuated the gain of WAT weight in KK- A(y) mice with increasing UCP1 expression compared with the control mice. The WAT weight of the mice fed 0.1% fucoxanthin and 6.9% fish oil was also significantly lower than that of the mice fed fucoxanthin alone. In addition, 0.2% fucoxanthin markedly decreased the blood glucose and plasma insulin concentrations in KK- A(y) mice. The mice fed with the combination diet of 0.1% fucoxanthin and fish oil also showed improvements similar to that of 0.2% fucoxanthin. Leptin and tumor necrosis factor (TNFalpha) mRNA expression in WAT were significantly down-regulated by 0.2% fucoxanthin. These results suggest that dietary fucoxanthin decreases the blood glucose and plasma insulin concentration of KK- A(y) along with down-regulating TNFalpha mRNA. In addition, the combination of fucoxanthin and fish oil is more effective for attenuating the weight gain of WAT than feeding with fucoxanthin alone.
Cytoprotective effect of fucoxanthin isolated from brown algae Sargassum siliquastrum against H2O2-induced cell damage
[J].
Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MARKs in lipopolysaccharide-induced RAW264.7 macrophages
[J].
Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway
[J].
Isolation of fucoxanthin from the rhizoid of Laminaria japonica Aresch
[J].
Extraction of fucoxanthin and polyphenol from Undaria pinnatifida using supercritical carbon dioxide with co-solvent
[J].
Supercritical carbon dioxide extraction of fucoxanthin from Undaria pinnatifida
[J].
Isolation of fucoxanthin from edible brown algae by microwave-assisted extraction coupled with high-speed countercurrent chromatography
[J].A rapid and efficient method for the separation and purification of fucoxanthin from edible brown algae by microwave-assisted extraction coupled with high-speed countercurrent chromatography was developed. The algae were first extracted using microwave-assisted extraction, then the dried extract was dissolved and directly introduced into the high-speed countercurrent chromatography system with a two-phase solvent system consisting of hexane-ethyl acetate-ethanol-water (5:5:6:4, v/v/v/v). The isolation was done in less than 75 min, and a total of 0.83 mg, 1.09 mg, and 0.20 mg fucoxanthin were obtained from 25.0 g fresh Laminaria japonica Aresch, 1.5 g dry Undaria pinnatifida (Harv) Sur, and 15.0 g dry Sargassum fusiforme (Harv) Setch, respectively. The purity of fucoxanthin determined by HPLC was over 90% and its structure was further identified by LC-ESI-MS and (1) H-NMR.© 2012 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim.
Pressurized liquid method for fucoxanthin extraction from Eisenia bicyclis(Kjellman)Setchell
[J].
Enzyme-assisted extraction of fucoxanthin and lipids containing polyunsaturated fatty acids from Undaria pinnatifida using diemethyl ether and ethanol
[J].
UPLC:An introduction and review
[J].
Arsenic speciation and fucoxanthin analysis from seaweed dietary supplements using LC-MS
[J].The study involves the analysis of total arsenic (As) in metallic form, and organic and inorganic As species from seaweeds and dietary supplements. The analysis provides data for dietary exposure estimates of inorganic species that are considered more toxic to humans than organic and total As. Total As was determined by acid digestion followed by inductively coupled plasma (ICP)-MS. To characterize the As species, solvent extraction with sonication and microwave extraction using various aqueous and aqueous/organic solvent mixtures were initially evaluated. The optimum As speciation method was determined to be water extraction followed by anion exchange HPLC coupled with ICP-MS. Optimization of chromatographic conditions led to baseline separation for six As species, including As acid, arsenous acid, monomethylarsonic acid, dimethylarsinic acid, arsenobetaine, and arsenocholine, in approximately 8 min using gradient elution. Detection limits for all six compounds were in the range of 10-15 ng/mL. The data presented here will be valuable for the QA of analytical method development and surveys of total As and As species in dietary supplements. The most abundant As species found were arsenite [As(III)] and arsenate [As(V)]. The sum of inorganic As species present in the dietary supplements ranged from 1.2 to 31 μg/day. In addition, the dietary supplements purported to contain fucoxanthin, a carotenoid having pharmacological activities, were analyzed using ultra-performance LC-UV/MS.
Apoptosis-Inducing effect of fucoxanthin on human leukemia cell line HL-60
[J].
营养盐亏缺与恢复对威氏海链藻(Thalassiosira weissflogii)生长和生化组成的影响
[J].
Phosphate-limitation mediated lipid production by Rhodosporidium toruloides
[J].
氮、磷、铁对三角褐指藻诱变株生长、总脂及脂肪酸的影响
[J].
为了优化微藻培养条件,采用单因子试验研究了不同氮浓度(5、10、15、20、25、30和35 mg·L<sup>-1</sup>)、不同磷浓度(0.5、1.0、1.5、2.0、2.5、3.0和3.5 mg·L<sup>-1</sup>)、不同铁源(FeCl<sub>3</sub>、FeC<sub>6</sub>H<sub>5</sub>O<sub>7</sub>和FeSO<sub>4</sub>)和铁浓度(0、0.10、0.25、0.50和1.00 mg·L<sup>-1</sup>)对三角褐指藻紫外诱变株MP2的影响。结果表明:氮、磷、铁对MP-2的生长、总脂含量和脂肪酸组成影响显著(P<0.05);MP-2生长最适氮浓度为20 mg·L<sup>-1</sup>,其生长速率K值和生物量分别为(0.384±0.004)和(0.25±0.01)g·L<sup>-1</sup>,氮浓度为30 mg·L<sup>-1</sup>时总脂积累量最高(26.51±1.96)%,氮浓度25 mg·L<sup>-1</sup>时PUFA积累最高[(37.78±0.35)%];MP-2生长最适磷浓度为2.5 mg·L<sup>-1</sup>,生长速率K值和生物量分别为(0.305±0.010)和(0.28±0.02)g·L<sup>-1</sup>,磷浓度1.0 mg·L<sup>-1</sup>时总脂积累量最高
氮限制对硅藻三角褐指藻光系统Ⅱ光化学反应的影响
[J].本文以硅藻三角褐指藻为实验材料,设置氮充足(882.35 μmol·L<sup>-1</sup>)和氮限制(40 μmol·L<sup>-1</sup>)两种营养盐供应水平进行培养,以探讨氮限制条件下三角褐指藻光系统Ⅱ(PSII)光化学反应及其对高光强的耐受性变化。结果表明:氮限制抑制三角褐指藻的生长,降低其叶绿素a含量,并加剧其受光抑制程度;通过分析快速叶绿素荧光诱导动力学曲线发现,氮限制使得电子从Q<sub>A</sub><sup>-</sup>向Q<sub>B</sub>的传递受阻,且高光强作用后,Q<sub>A</sub><sup>-</sup>的瞬时积累进一步增加;此外,氮限制条件下,活性反应中心部分关闭,藻细胞单位反应中心吸收的能量(ABS/RC)、捕获的用于还原Q<sub>A</sub>的能量(TR<sub>o</sub>/RC)以及耗散的能量(DI<sub>o</sub>/RC)均明显升高;而高光的耦合作用使得PSII反应中心进一步失活;氮限制影响三角褐指藻PSII光化学反应,进而降低其对高光的耐受能力。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}