


枯草芽孢杆菌(Bacillus subtilis),隶属于厚壁菌门(Firmicutes)、芽孢杆菌科(Bacillaceae)、芽孢杆菌属(Bacillus),具有耐高温、耐酸、耐碱、稳定性高的特性。养殖水体中投放适量的枯草芽孢杆菌可以降低水体亚硝酸盐含量,维持水体pH及溶解氧含量,起到净化水质的作用[1-2];饲料中添加枯草芽孢杆菌可提高罗非鱼(Oreochromis niloticus)的生长速度、消化酶活性以及先天性免疫反应和抗病力[3];口服芽孢杆菌可以提高南亚野鲮(Labeo rohita)的呼吸暴发活性以及先天性免疫反应[4]。作为水产养殖上应用较广泛、极具潜力的有益菌种之一[5],枯草芽孢杆菌可以抑制有害菌生长,调整肠道微生态菌群[6],提高消化酶,促进脂肪代谢,降低饵料系数,提高鱼体对营养物质的吸收[7-8]。
1 材料与方法
1.1 实验材料
于2021年5月20日— 6月31月在福建省淡水水产研究所榕桥中试基地开展实验,水源为水库水。实验用鱼均由福建省淡水水产研究所榕桥中试基地提供,健康黑脊倒刺鲃(记为Sc)平均体质量和平均体长分别为(11.0±0.29)g和(14.2±0.74)cm;半刺厚唇鱼(记为Ah)平均体质量和平均体长分别为(6.8±0.56)g和(3.5±0.36)cm;鲢(记为Hm)平均体质量和平均体长分别为(15.8±0.25)g和(33.3±2.8)cm。枯草芽孢杆菌由北京渔经生物技术有限公司提供,芽孢含量达1012 CFU/g。投喂粗蛋白含量为31%的2#淡水鱼膨化颗粒配合饲料,配合饲料购自天马科技集团股份有限公司。
1.2 鱼种放养与日常管理
实验容器为150 L玻璃纤维桶,每桶装水120 L,混养上述3种实验鱼各20尾,连续充气,暂养7 d,待稳定后开始实验,实验设置投放芽孢杆菌的实验组(Experiment group,E)和不投放芽孢杆菌的对照组(Control group, C)。实验组按照1.0 g/m3枯草芽孢杆菌粉剂的水体浓度进行泼洒,使用前以枯草芽孢杆菌∶红糖∶水=2∶1∶50比例进行洁净水活化2 h。实验期间上午、下午各投喂饲料1次,日投饵量约为3种鱼体总体质量的1.5%,日换水1次,换水量为40%,实验组换水后,仍按照1.0 g/m3枯草芽孢杆菌剂量并活化2 h后再泼洒,将水体中枯草芽孢杆菌含量维持在105 cfu/mL,对照组不投放芽孢杆菌。每天早上08:00,在投喂前检测养殖水体水温、pH值、氨氮和亚硝酸盐含量,检测仪器为格蓝普智能多参数水质检测仪(型号GLP-T100)。
1.3 肠道菌群 16S rDNA 测序
实验后31 d,随机采集实验组和对照组养殖桶黑脊倒刺鲃、半刺厚唇鱼和鲢各2尾,合计12尾,分别取鱼体的前肠(Foregut,F)、中肠(Midgut,M)和后肠(Hindgut,H)内容物,黑脊倒刺鲃实验组和对照组前、中、后肠分别命名为Sc_F31E、Sc_F31C、Sc_M31E、Sc_M31C和Sc_H31E、Sc_H31C;半刺厚唇鱼实验组和对照组前、中、后肠分别命名Ah_F31E、Ah_F31C、Ah_M31E、Ah_M31C和Ah_H31E、Ah_H31C;鲢实验组和对照组前、中、后肠分别命名为Hm_F31E、Hm_F31C、Hm_M31E、Hm_M31C和Hm_H31E、Hm_H31C(以下同此)。液氮速冻后-80 ℃冰箱保存,送至杭州联川生物技术股份有限公司进行16S rDNA 测序分析。扩增引物为341F:5’CCTACGGGNGGCWGCAG-3’;805R:5’-GACTACHVGGGTATCTAATCC-3’,测序平台为NovaSeq 6000测序仪,测序策略为PE250双端测序。
1.4 生物信息学分析
1.4.1 数据处理
对测序reads进行质控,获得高质量的Clean data。通过DADA2(Divisive amplicon denoising algorithm)[12]进行长度过滤和去噪,以100%相似度进行单元聚类获得ASV (Amplicon sequence variant)特征序列和丰度表格。
1.4.2 Alpha多样性分析
基于得到的ASV特征序列和丰度表格进行Alpha多样性分析。用Chao1指数和Observed species指数评估群落中所包含物种的数目;用Shannon-Weiner指数和Simpson指数来评估群落的多样性。评判标准为Chao1指数和Observed species指数越大,物种数目越多;Shannon-Weiner指数越大,多样性越高;Simpson指数越小,群落多样性越高。
1.4.3 PCoA分析
主坐标分析PCoA是基于距离矩阵,通过一系列的特征值和特征向量排序后,以可视化二维空间重新排列样品,最大化展示样品之间的关系信息[13]。PCoA分析中样品点之间的距离越近,说明样品之间物种组成结构越相似。
1.4.4 功能预测分析
运用PICRUST软件与Greengene数据库进行物种组成信息比对[14],计算相应KEGG的信息及丰度,得到相应代谢通路,在95%置信度区间内对各实验组和对照组进行t-test和P-value检验,分析其在功能上的差异。
2 结果与分析
2.1 水质指标变化
实验期间水温为23.5~27.5 ℃,其中实验组水体芽孢杆菌含量维持在(1.6~2.5)×1
图1
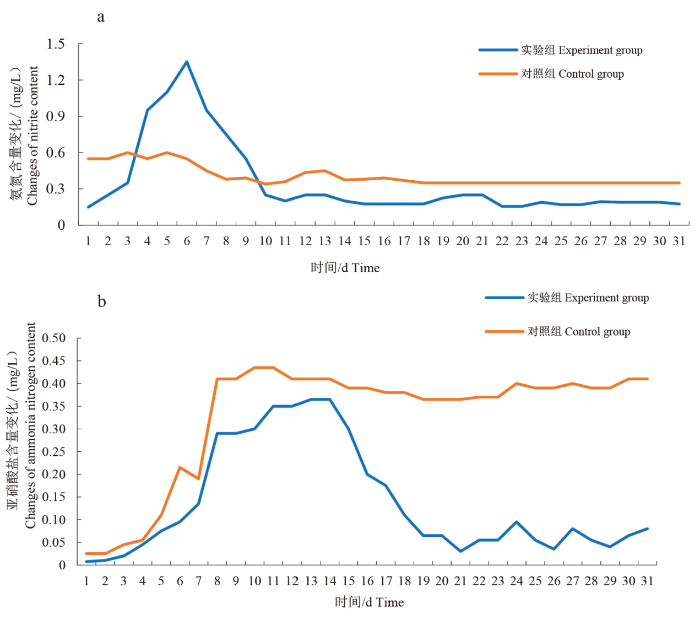
图1
实验期间水体氨氮(a)、亚硝酸盐(b)含量变化
Fig.1
The content variation of ammonia nitrogen (a) and nitrite (b) in the test duration
2.2 测序样本序列信息
测序36个样本下机数据量为974.46 M,平均测序数据量为27.07 M。数据共产生2 349 864条raw tags、Clean tags,每个样本平均有65 274条raw tags、Clean tags,数据有效性为100%,99.91%序列分布在400~500 bp之间,平均GC含量为54.87%。测序样本覆盖率均大于98%,说明测序数据量足以代表样品的细菌群落。
2.3 微生物群落Alpha多样性分析
实验组黑脊倒刺鲃前肠、半刺厚唇鱼后肠Observed specie指数上升,上升比例分别为26%和31%;Chao1指数上升,上升比例分别为27%和19%。其他鱼体肠道部位Observed species和Chao1指数均下降,下降比例分别为12%~94%和15%~94%。黑脊倒刺鲃中肠、半刺厚唇鱼后肠Simpson指数上升,上升比例分别为22%和68%; Shannon-Weiner指数上升,上升比例分别为15%和155%。其他鱼体肠道部位Simpson和Shannon-Weiner指数均下降,下降比例分别为1%~34%和3%~72%(图2)。
图2
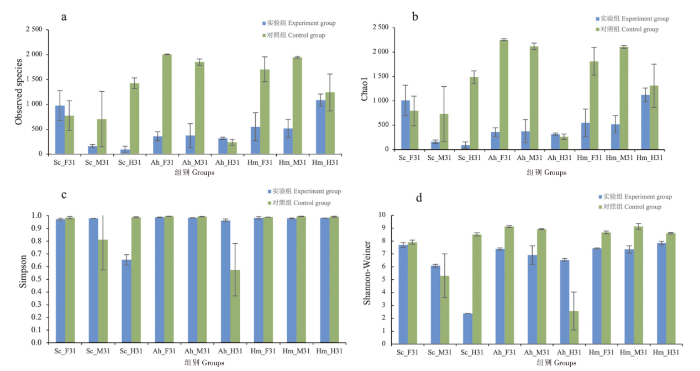
图2
三种鱼体不同肠道部位微生物群落Alpha 多样性变化
注:Sc_F31.黑脊倒刺鲃前肠;Sc_M31.黑脊倒刺鲃中肠;Sc_H31.黑脊倒刺鲃后肠;Ah_F31.半刺厚唇鱼前肠;Ah_M31.半刺厚唇鱼中肠;Ah_H31.半刺厚唇鱼后肠;Hm_F31.鲢前肠;Hm_M31.鲢中肠;Hm_H31.鲢后肠。以下同此。
Fig.2
Changes of Alpha diversity of microbial community in different intestinal sites of three fish species
Notes: Sc_F31 was the forgut of S.caldwelli; Sc_M31 was the midgut of S.caldwelli; Sc_H31 was the hindgut of S.caldwelli; Ah_F31 was the forgut of A.hemispinus; Ah_M31 was the midgut of A.hemispinus;Ah_H31 was the hindgut of A.hemispinus; Hm_F31 was the forgut of H.molitri; Hm_F31 was the midgut of H.molitri; Hm_F31 was the hindgut of H.molitri.The same as below.
整体而言,3种鱼类各肠道部位实验组Observed species和Chao1指数下降,Shannon-Weiner和Simpson指数下降,细菌丰富度降低。
2.4 PCoA分析
黑脊倒刺鲃、半刺厚唇鱼和鲢PCoA分析最主要特征值PCoA1和PCoA2对样本的差异的贡献值分别合计达到82.35%、83.02%和79.94%,可以代表大部分变量信息。除黑脊倒刺鲃前肠和鲢后肠外,3种鱼类其他不同肠道部位中,实验组与对照组各自区分聚类,可用于后续信息分析(图3)。
图3
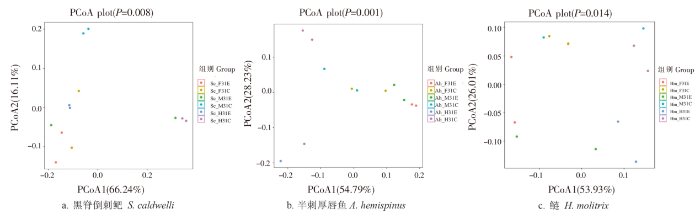
图3
三种鱼类前、中、后肠道菌群PCoA 分析
注:组别编号最后一个字母E表示实验组,C表示对照组。以下同此。
Fig. 3
PCoA analysis of gut microbiota in the foregut, midgut and hindgut of the three fishes
Notes:The last letter of the group number E represented the experimental group, and C represented the control group.The same as below.
2.5 基于门水平的微生物群落结构分析
如图4所示,黑脊倒刺鲃前肠对照组和实验组的优势菌门为变形菌门、浮霉菌门和放线菌门,梭杆菌门和厚壁菌门分别是两组区别优势菌门,优势菌门合计占比分别为71.97%和82.89%。中肠对照组和实验组的优势菌门为变形菌门和厚壁菌门,浮霉菌门、梭杆菌门和放线菌门、拟杆菌门分别是两组区别优势菌门,优势菌门合计占比分别为85.69%和86.69%。后肠实验组的优势菌门为变形菌门,占比高达98.47%,对照组共有优势菌门为变形菌门,区别优势菌门为浮霉菌门、放线菌门和拟杆菌门,优势菌门合计占比为71.40%。与对照组相比,浮霉菌门和梭杆菌门在实验组前、中、后肠的丰度均下降;变形菌门在前、后肠的丰度上升,在中肠下降;放线菌门和厚壁菌门在前、中肠的丰度上升,在后肠下降。
图4
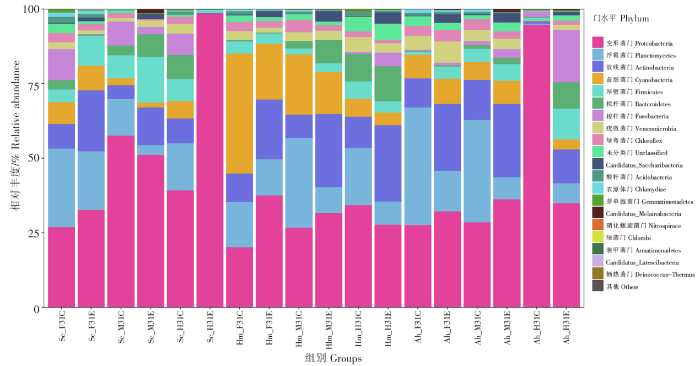
图4
三种鱼类肠道菌群门水平差异分析
Fig.4
Variation analysis of gut microbiota at phylum level among three fish species
半刺厚唇鱼前肠实验组、对照组和中肠实验组、对照组的优势菌门为变形菌门、浮霉菌门、放线菌门和蓝细菌门,合计占比分别为76.6%、84.63%和75.96%、82.24%。后肠对照组优势菌门为变形菌门和梭杆菌门,合计占比96.57%,实验组共有优势菌门为变形菌门,区别优势菌门为梭杆菌门、放线菌门和厚壁菌门,合计占比为74.25%。实验组放线菌门、蓝细菌门、厚壁菌门、梭杆菌门在前、中、后肠的丰度均上升,变形菌门在前肠的丰度上升,在中、后肠的丰度下降;浮霉菌门在前、中肠的丰度上升,在后肠的丰度下降。
鲢前、中、后肠道实验组和对照组的优势菌门一致。前肠实验组、对照组和中肠实验组、对照组的优势菌群均为变形菌门、浮霉菌门、放线菌门和蓝细菌门,合计占比分别为88.33%、85.05%和78.81%、84.76%。鲢后肠实验组和对照组的优势菌群为变形菌门、浮霉菌门、放线菌门和拟杆菌门,合计占比分别为72.78%和73.27%。放线菌门在前、中、后肠道的丰度上升,蓝细菌门、浮霉菌门在前、中、后肠道的丰度下降;变形菌门在前、中肠的丰度上升,在后肠的丰度下降;拟杆菌门在中、后肠的丰度上升,在前肠的丰度下降。
3种鱼类共有的优势菌门均为变形菌门,其他优势菌门在各组中均有一定的差异。整体而言,实验组放线菌门在3种鱼类各个肠道部位的丰度均上升,浮霉菌门在3种鱼类各个肠道部位的丰度均下降。3种鱼的前肠变形菌门和放线菌门丰度均上升,中肠道放线菌门丰度均上升,后肠道浮霉菌门丰度均下降。
2.6 基于属水平的微生物群落结构分析
如图5所示,黑脊倒刺鲃前肠对照组和实验组共有优势菌属为浮霉菌门未分类属和蓝细菌门未分类属,鲸杆菌属和Demequina属是各自区别优势菌属,各优势菌属合计占比分别为30.83%和28.89%。气单胞属为中肠对照组第一优势菌属,占比为45.14%,其他优势菌属为鲸杆菌属、厚壁菌门未分类属;实验组优势菌属为厚壁菌门未分类属、浮霉菌门未分类属和芽孢杆菌属,合计占比为18.04%。后肠对照组优势菌属为β-变形菌纲未分类属、鲸杆菌属、浮霉菌门未分类属,合计占比为23.09%;实验组优势菌属主要为气单胞属,占比为93.54%。与对照组相比,实验组浮霉菌门未分类属、鲸杆菌属、弧菌属、出芽菌属在前、中、后肠的丰度均下降,芽孢杆菌属在前、中、后肠的丰度均上升。
图5
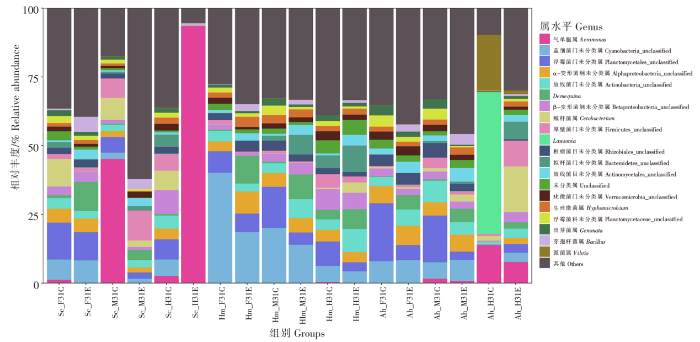
图5
三种鱼类肠道菌群属水平差异分析
Fig.5
Variation analysis of gut microbiota at genus level among three fish species
蓝细菌门未分类属和α-变形菌纲未分类属是半刺厚唇鱼前肠对照组和实验组的优势菌属,浮霉菌门未分类属和放线菌门未分类属分别是两组的差别优势菌属,各优势菌属合计占比分别为35.30%和21.25%。蓝细菌门未分类属是中肠对照组和实验组的共有优势菌属,浮霉菌门未分类属、根瘤菌目未分类属和α-变形菌纲未分类属、Demequina属分别是两组的差别优势菌属,合计占比分别为28.28%和18.73%。Lawsonia属是后肠对照组第一优势菌属,占比为51.49%,第二、三优势菌属为弧菌属、气单胞属,占比分别为20.23%和13.89%;后肠实验组优势菌属为鲸杆菌属、厚壁杆菌门未分类属和气单胞属,合计占比为33.69%。蓝细菌门未分类属、Demequina属、芽孢杆菌属在前、中、后肠道的丰度均上升,气单胞属、弧菌属和Lawsonia属在前、中、后肠道的丰度均下降。
鲢前肠对照组和实验组共有优势菌属为蓝细菌门未分类属,浮霉菌门未分类属、放线菌门未分类属和α-变形菌纲未分类属、Demequina属分别是两组的区别优势菌属,合计占比分别为51.73%和36.62%。鲢中肠对照组和实验组共有优势菌属为蓝细菌门未分类属,浮霉菌门未分类属、α-变形菌纲未分类属和放线菌门未分类属和Demequina属是两组的区别优势菌属,合计占比分别为40.04%和29.80%。鲢后肠对照组和实验组共有优势菌属为β-变形菌纲未分类属,蓝细菌门未分类属、浮霉菌门未分类属和放线菌门未分类属、Demequina属是两组的区别优势菌属,合计占比分别为22.26%和21.64%。Demequina属和芽孢杆菌属在前、中、后肠的丰度均上升,气单胞属、蓝细菌门未分类属、浮霉菌门未分类属和弧菌属在前、中、后肠的丰度均下降。
3种鱼类肠道菌群以及同种鱼类不同肠道在属水平优势菌群差异较大,各肠道样本菌属比较丰富,且丰度较为均一。整体而言,气单胞属、弧菌属在3种鱼类各肠道部位的丰度均下降,芽孢杆菌属在3种鱼类各肠道部位的丰度均上升。
2.7 三种鱼类微生物菌落功能差异分析
黑脊倒刺鲃中肠外源性物质生物降解与代谢和后肠酶家族、代谢、碳水化合物代谢以及氨基酸代谢提高,前肠多糖生物合成与代谢、后肠外源性物质生物降解与代谢、萜类和多酮类化合物代谢、酯类代谢降低。半刺厚唇鱼前肠和中肠酶家族降低,中肠的其他氨基酸代谢,后肠脂类代谢、萜类和多酮类化合物代谢提高。鲢前肠碳水化合物代谢,中肠其他氨基酸代谢,后肠其他次生代谢物生物合成、萜类和多酮类化合物代谢提高(表1)。
表1 三种鱼类不同肠道部位菌群KEGG信号通路相对丰度差异
Tab.1
| 肠道部位 Intestinal sites | 途径 Pathways | 相对丰度/% Relative abundance | ||
|---|---|---|---|---|
| 实验组 Experiment group | 对照组 Control group | |||
| 黑脊倒刺鲃 S.caldwelli | 前肠 | 多糖生物合成与代谢 | 1.871±0.022 | 2.183±0.003 |
| 中肠 | 外源性物质生物降解与代谢 | 2.920±0.105 | 1.874±0.194 | |
| 后肠 | 外源性物质生物降解与代谢 | 1.763±0.068 | 2.102±0.058 | |
| 信号转导 | 3.133±0.026 | 2.133±0.052 | ||
| 萜类和多酮类化合物代谢 | 1.126±0.009 | 1.634±0.028 | ||
| 代谢 | 3.064±0.034 | 2.067±0.020 | ||
| 脂类代谢 | 2.916±0.016 | 3.568±0.044 | ||
| 酶家族 | 2.435±0.001 | 2.184±0.012 | ||
| 细胞过程和信号转导 | 5.438±0.033 | 3.664±0.068 | ||
| 细胞运动 | 5.380±0.151 | 2.889±0.154 | ||
| 碳水化合物代谢 | 9.687±0.207 | 7.973±0.043 | ||
| 氨基酸代谢 | 9.701±0.084 | 7.852±0.076 | ||
| 半刺厚唇鱼 A.hemispinus | 前肠 | 酶家族 | 2.066±0.006 | 2.196±0.007 |
| 折叠、分类和降解 | 2.757±0.026 | 2.968±0.016 | ||
| 膜运输 | 12.45±0.083 | 11.040±0.035 | ||
| 中肠 | 酶家族 | 2.053±0.007 | 2.170±0.014 | |
| 其他氨基酸代谢 | 1.883±0.012 | 1.743±0.010 | ||
| 后肠 | 细胞运动 | 2.581±0.011 | 5.214±0.207 | |
| 遗传信息处理 | 2.562±0.028 | 3.009±0.046 | ||
| 脂类代谢 | 3.293±0.107 | 2.680±0.144 | ||
| 萜类和多酮类化合物代谢 | 1.611±0.005 | 1.169±0.032 | ||
| 鲢 H.molitrix | 前肠 | 碳水化合物代谢 | 9.583±0.024 | 9.138±0.045 |
| 遗传信息处理 | 2.501±0.004 | 2.699±0.002 | ||
| 转录 | 2.247±0.029 | 2.000±0.035 | ||
| 中肠 | 其他氨基酸代谢 | 1.871±0.012 | 1.745±0.011 | |
| 后肠 | 其他次生代谢物生物合成 | 1.021±0.014 | 0.960±0.011 | |
| 萜类和多酮类化合物代谢 | 1.696±0.007 | 1.662±0.005 | ||
3种鱼类实验组非代谢信号途径中,黑脊倒刺鲃后肠信号转导、细胞过程和信号转导以及细胞运动提高;半刺厚唇鱼前肠膜运输信号通路提高,后肠细胞运动和遗传信息处理水平降低;鲢前肠遗传信息处理和转录水平降低。
3 讨论
枯草芽孢杆菌影响水体环境和鱼体肠道需要一定的时间,一般为2~4周[1,10]。因本研究中使用的3种鱼类习性不同,为使枯草芽孢杆菌在3种鱼体中能够稳定定植,研究周期延长至31 d,结果显示投放枯草芽孢杆菌株后,水体pH变化不大,这与杭小英等[15]、刘慧玲等[16]的研究报道一致。国内研究大量报道了枯草芽孢杆菌能够降低水体氨氮含量、亚硝酸盐含量,起到净化水质的作用[1,10,15],主要是因为枯草芽孢杆菌可通过自养反硝化和自养亚硝化而降低水中氨氮、亚硝酸态氮的含量[17]。本研究初期投放枯草芽孢杆菌时养殖水体氨氮含量上升,可能是由于枯草芽孢杆菌进入水体,消耗了大量的有机氮,从而造成氨氮短时间的积累[2]。水体中保持一定浓度的枯草芽孢杆菌后,3种鱼类实验组各肠道部位菌群整体种类减少,丰富度下降。研究证明,水产动物成鱼的肠道菌群Alpha多样性一般低于幼鱼,低水平的Alpha多样性有利于肠道菌群的稳定[18⇓-20]。
枯草芽孢杆菌通过与致病菌竞争生存空间、定居部位以及营养物质等,可以有效地抑制有害菌的生长和繁殖,改善鱼类肠道内微生物群落的组成。同时,枯草芽孢杆菌可以分泌产生酸性物质、细菌素等抑制或杀灭有害菌,调整肠道微生态平衡[21]。有研究报道,日粮中添加该菌可以减少草鱼(Ctenopharyngodon idella)肠道菌群中假单胞菌和和黄杆菌等条件致病菌的增殖[22];水体中投放该菌可以抑制克氏原鳌虾(Procambarus clarkii)大肠菌群和金黄色葡萄球菌等有害致病菌的增殖[23]。本研究水体中的枯草芽孢杆菌抑制了3种鱼类中肠道菌群气单胞属、弧菌属的增殖。由此可见,枯草芽孢杆菌作为水体或饲料添加剂,均能改善鱼体肠道菌群,抑制有害病原菌的增殖。本研究中气单胞属在黑脊倒刺鲃实验组大量繁殖的原因有待进一步研究。有报道称气单胞属也是黄颡鱼(Pelteobagrus fulvidraco)的优势菌属,其可能对黄颡鱼的免疫系统发育起到一定的促进作用[24]。芽孢杆菌作为水产常用的微生态制剂,可以合成益菌物质,抑制细菌、真菌和病毒生长,同时形成生物大分子降解酶,促进鱼体消化[21]。本研究中芽孢杆菌属在3种鱼类各肠道部位的丰度均上升,说明枯草芽孢杆菌可以促进肠道有益菌的生长与定植。
大量文献[25⇓-27]报道枯草芽孢杆菌可以增强鱼类代谢功能、提高脂肪利用率、促进鱼类生长。使用枯草芽孢杆菌投喂杂交鲟[西伯利亚鲟Acipenser baerii♀×施氏鲟A.schrenckii♂],可显著降低饵料系数,增强鱼体对血糖和脂肪的利用和转化能力[25];投喂草鱼枯草芽孢杆菌,其肝脏油及脂肪含量显著下降,脂蛋白脂酶及脂肪甘油三酯脂肪酶的表达水平上调[26],跟脂质代谢通路相关的基因表达几乎上调[27]。本研究中枯草芽孢杆菌加强了鲢的肠道菌群代谢功能,对其他2种鱼类则效果不佳,产生这一差异可能是因为鲢是滤食性鱼类,具海绵状的鳃耙和较长的肠道,能够有效过滤吸收细菌[28]。胡保同[29]在鲢肠道前端找到被黏液包裹的细菌凝聚体;朱学宝[30]以细菌絮凝体作为直接饵料饲养鲢,13 d后鲢鱼增重37.32%。在自然湖泊中,鲢摄取的细菌数量可占食物总量的1/3。因此相较于半刺厚唇鱼和黑脊倒刺鲃,鲢更容易摄食和吸收水体中的枯草芽孢杆菌。
国内外关于水体投放枯草芽孢杆菌对鱼类肠道菌群影响的报道较少,本研究选取生活于不同水层、不同食性的鱼类作为研究对象,探讨该菌对3种鱼类不同肠道部位菌群和代谢的变化。然而由于样本取样数目的限制导致相关统计学分析无法展开,因此本研究只能基于已有的数据,分析不同肠道门、属和功能的差异变化。由于不同鱼类以及同一鱼类不同肠道菌群差异较大,本文只能简单整合、比较同一鱼类和同一肠道部位共有的菌群变化,以期为后续相关水体投放枯草芽孢杆菌对鱼类肠道菌群影响的研究提供一定的数据支撑和借鉴价值。
参考文献
Dietary administration of Bacillus subtilis HAINUP40 enhances growth, digestive enzyme activities, innate immune responses and disease resistance of tilapia, Oreochromis niloticus
[J].
Enhanced innate immune parameters in Labeo rohita (Ham.) following oral administration of Bacillus subtilis
[J].
Probiotic applications for rainbow trout (Oncorhynchus mykiss Walbaum):Ⅰ. Effects on growth performance, feed utilization, intestinal microbiota and related health criteria
[J].
四种非营养性饲料添加剂对澳洲淡水龙虾消化酶活力、免疫功能及离体消化率的影响
[J].
探讨4种非营养性饲料添加剂对澳洲淡水龙虾(Cherax quadricarinatus)肝胰腺消化酶活力、免疫功能及离体消化率的影响,筛选出适宜的饲料添加剂,为非营养性添加剂在饲料中的应用和研究提供理论依据。分别以柠檬酸(S-Ⅰ)、复合酶制剂(S-Ⅱ)、植物乳杆菌加枯草芽孢杆菌(S-Ⅲ)、根瘤菌加枯草芽孢杆菌(S-Ⅳ)作为非营养性添加剂,配制4种等氮等脂等能饲料,喂养澳洲淡水龙虾14 d后,取样测定其肝胰腺消化酶(胰蛋白酶、脂肪酶和淀粉酶)活力、免疫功能[碱性磷酸酶(AKP)、酸性磷酸酶(ACP)、超氧化物歧化酶(SOD)活力和总抗氧化能力(T-AOC)]及离体消化率(干物质消化率、蛋白质离体消化率和总氨基酸转化效率)等相关指标。结果表明,S-Ⅰ可以显著提高澳洲淡水龙虾肝胰腺的淀粉酶、AKP和SOD活力(P<0. 05),S-Ⅱ可以显著提高其淀粉酶和胰蛋白酶活力以及免疫功能(P<0. 05);而S-Ⅲ和S-Ⅳ在消化酶活力和免疫功能上均显著提高(P<0. 05)。在4种非营养性饲料添加剂对澳洲淡水龙虾离体消化率的影响中,S-Ⅳ的干物质消化率、蛋白质离体消化率和总氨基酸转化效率均值最高。S-Ⅲ和S-Ⅳ对消化酶活力、免疫功能及离体消化率的影响比S-Ⅰ和S-Ⅱ的高,并且S-Ⅳ的离体消化率比S-Ⅲ高,因此在这4种非营养性添加剂中优先选择S-Ⅳ。
等珍珠龙胆石斑鱼肠道枯草芽孢杆菌的分离鉴定及产酶能力分析
[J].从健康珍珠龙胆石斑鱼肠道中分离纯化获得4株疑似枯草芽孢杆菌(Bacillus subtilis)菌株(B1、B2、B3和B4),并进行了菌落形态、菌株理化性质、16S rDNA序列和产酶能力分析等方面的研究。结果表明,在理化性质方面,4株菌的试验结果均符合枯草芽孢杆菌的生化特性,并结合菌落形态,初步证明4株分离菌可能均为枯草芽孢杆菌;同时,基因序列比对结果显示,4株菌的16S rDNA序列与已知的枯草芽孢杆菌序列相似性≥99%,最终鉴定均为枯草芽孢杆菌。经过产酶能力的试验研究发现,4株菌都具有产纤维素酶、淀粉酶和蛋白酶的能力,且B3菌株同其他3株菌相比,其产纤维素酶、淀粉酶和蛋白酶的能力均较强。因此,本试验选取B3作为具有益生菌潜力的菌株。
Control of pathogenic Vibrio spp. by Bacillus subtilis BT23, a possible probiotic treatment for black tiger shrimp Penaeus monodon
[J].The present study evaluated the in vitro and in vivo antagonistic effect of Bacillus against the pathogenic vibrios.Cell-free extracts of Bacillus subtilis BT23 showed greater inhibitory effects against the growth of Vibrio harveyi isolated by agar antagonism assay from Penaeus monodon with black gill disease. The probiotic effect of Bacillus was tested by exposing shrimp to B. subtilis BT23 at a density of 106-108 cfu ml-1 for 6 d before a challenge with V. harveyi at 103-104 cfu ml-1 for 1 h infection. The combined results of long- and short-term probiotic treatment of B. subtilis BT23 showed a 90% reduction in accumulated mortality.This study reports that pathogenic vibrios were controlled by Bacillus under in vitro and in vivo conditions.Results indicated that probiotic treatment offers a promising alternative to the use of antibiotics in shrimp aquaculture.
Effects of Bacillus subtilis on the water quality, stress tolerance, digestive enzymes, growth performance, immune gene expression, and disease resistance of white shrimp (Litopenaeus vannamei) during the early hatchery period
[J].
DADA2: high-resolution sample inference from illumina amplicon data
[J].We present the open-source software package DADA2 for modeling and correcting Illumina-sequenced amplicon errors (https://github.com/benjjneb/dada2). DADA2 infers sample sequences exactly and resolves differences of as little as 1 nucleotide. In several mock communities, DADA2 identified more real variants and output fewer spurious sequences than other methods. We applied DADA2 to vaginal samples from a cohort of pregnant women, revealing a diversity of previously undetected Lactobacillus crispatus variants.
Some distance properties of latent root and vector methods used in multivariate analysis
[J].
Pre-dictive functional profiling of microbial communities using 16S rRNA marker gene sequences
[J].Profiling phylogenetic marker genes, such as the 16S rRNA gene, is a key tool for studies of microbial communities but does not provide direct evidence of a community's functional capabilities. Here we describe PICRUSt (phylogenetic investigation of communities by reconstruction of unobserved states), a computational approach to predict the functional composition of a metagenome using marker gene data and a database of reference genomes. PICRUSt uses an extended ancestral-state reconstruction algorithm to predict which gene families are present and then combines gene families to estimate the composite metagenome. Using 16S information, PICRUSt recaptures key findings from the Human Microbiome Project and accurately predicts the abundance of gene families in host-associated and environmental communities, with quantifiable uncertainty. Our results demonstrate that phylogeny and function are sufficiently linked that this 'predictive metagenomic' approach should provide useful insights into the thousands of uncultivated microbial communities for which only marker gene surveys are currently available.
Ontogenetic differences in dietary fat influence microbiota assembly in the zebrafish gut
[J].
Environmental filtering decreases with fish development for the assembly of gut microbiota
[J].Gut microbiota typically occupy habitats with definable limits/borders that are comparable to oceanic islands. The gut therefore can be regarded as an 'island' for the assembly of microbial communities within the 'sea' of surrounding environments. This study aims to reveal the ecological mechanisms that govern microbiota in the fish gut 'island' ecosystem. Taxonomic compositions, phylogenetic diversity, and community turnover across host development were analyzed via the high-throughput sequencing of 16S rRNA gene amplicons. The results indicate that the Shannon diversity of gut microbiota in the three examined freshwater fish species all significantly decreased with host development, and the dominant bacterial taxa also changed significantly during host development. Null model and phylogenetic-based mean nearest taxon distance (MNTD) analyses suggest that host gut environmental filtering led to the assembly of microbial communities in the fish gut 'island'. However, the phylogenetic clustering of local communities and deterministic processes that governed community turnover became less distinct as the fish developed. The observed mechanisms that shaped fish gut microbiota seemed to be mainly shaped by the gut environment and by some other selective changes accompanying the host development process. These findings greatly enhance our understanding of stage-specific community assembly patterns in the fish gut ecosystem.© 2016 The Authors Environmental Microbiology published by Society for Applied Microbiology and John Wiley & Sons Ltd.
The composition of the zebrafish intestinal microbial community varies across development
[J].The assembly of resident microbial communities is an important event in animal development; however, the extent to which this process mirrors the developmental programs of host tissues is unknown. Here we surveyed the intestinal bacteria at key developmental time points in a sibling group of 135 individuals of a model vertebrate, the zebrafish (Danio rerio). Our survey revealed stage-specific signatures in the intestinal microbiota and extensive interindividual variation, even within the same developmental stage. Microbial community shifts were apparent during periods of constant diet and environmental conditions, as well as in concert with dietary and environmental change. Interindividual variation in the intestinal microbiota increased with age, as did the difference between the intestinal microbiota and microbes in the surrounding environment. Our results indicate that zebrafish intestinal microbiota assemble into distinct communities throughout development, and that these communities are increasingly different from the surrounding environment and from one another.
Effects of dietary administration of Shewanella xiamenensis A-1,Aeromonas veronii A-7,and Bacillus subtilis, single or combined, on the grass carp (Ctenopharyngodon idella) intestinal microbiota
[J].
Comparative study on gastrointestinal microbiota of eight fish species with different feeding habits
[J].To reveal the effects of fish genotype, feeding habits and serum physiological index on the composition of gastrointestinal microbiota, eight fish species with four different feeding habits were investigated.The V1 to V3 regions of 16S rRNA gene were analysed by high-throughput sequencing (454 platform) to compare the gut microbiota of different fish species. A total of 551 995 high-quality sequences with an average length of 463 bp were obtained from the 48 samples. No significant difference was observed among the detected sequences obtained from fishes with different feeding habits (One-way anova, F = 1·003, P = 0·400), but the number of OTUs among different feeding habits was significantly different (One-way anova, F = 7·564, P < 0·001). Additionally, significant correlations were detected between the fish genotype and microbial composition (partial Mantel test, all P values = 0·001) in the stomach, foregut and hindgut. Moreover, different core intestinal microbiota was also noticed in the eight fish species with different feeding habits.Feeding habits and genotype clearly affected the gastrointestinal microbiota of fish. Moreover, the evolutionary process shaped the serum physiological indexes of fish.This study provided much important information for developing commercial fish feeds.© 2014 The Society for Applied Microbiology.
Bacillus subtilis supplementation in a high-fat diet modulates the gut microbiota and ameliorates hepatic lipid accumulation in grass carp (Ctenopharyngodon idella)
[J].To study the effects of Bacillus subtilis supplementation in a high-fat diet on the gut microbiota and nonalcoholic fatty liver disease in grass carp (Ctenopharyngodon idella), juveniles (60 ± 5 g) were fed three diets: (a) a control diet (CON), (b) a high-fat diet (HFD) and (c) a high-fat diet supplemented with B. subtilis (HFD + BS). After 8 weeks of feeding, fish growth, serum biochemical indices and total liver lipid content were measured, and gut microbiota analysis was performed using the MiSeq250 high-throughput sequencing platform. The results of this study showed that B. subtilis could improve growth and blood serum indices and reduce lipid deposition in the fish liver, preventing fatty liver disease. A grass carp model of fatty liver induced by a high-fat diet was successfully established. Moreover, B. subtilis altered the intestinal microbiota of HFD-fed grass carp, making it more similar to that of the control group. This study revealed the important effects of B. subtilis on grass carp with fatty liver induced by a high-fat diet and provides the foundation for the application of probiotics in grass carp farming.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}