


海带(Saccharina japonica)位居我国三大经济海藻之首,2020年全国海带年产量约为165.16×104 t[1]。海带苗种生产目前采用两种方法:孢子体育苗法和配子体育苗法。孢子体育苗法在海带育苗生产中被普遍使用,且大大促进了我国海带养殖产业的发展,但孢子体采苗技术存在大量海带藻体植株作为亲本培育苗种易导致种质退化、苗种培育受季节严格限制、育苗生产期较长、制冷成本高等不足[2]。配子体育苗法的亲本种质单一纯净,且不受季节限制[3],可有效避免孢子体采苗法的不足,但存在一个较大技术问题,即配子体苗种容易受敌害生物底栖硅藻污染,导致苗种质量下降。因此,目前配子体育苗法在条件易控的试验研究中应用较多,在海带育苗生产中应用较少。
海带配子体育苗分为两个大的阶段:一是配子体扩增培养阶段,二是配子体苗种培育阶段。针对配子体扩增培养条件已有较多的研究,包括光照强度和光照周期[4⇓-6]、温度[7⇓⇓⇓⇓-12]、培育水体的消毒方式[13]等,目前配子体扩增培养的技术已较为成熟,可有效保障配子体育苗生产。而对配子体苗种培育阶段的研究相对较少。张壮志等[14]研究了二次育苗对海带配子体苗种质量的影响,而对敌害生物底栖硅藻污染防治的研究尚未见报道。底栖硅藻污染问题在海带孢子体育苗生产中同样存在,但海带种菜释放的游孢子可以牢固黏附到苗种绳上,并逐步发育成胚孢子、配子体、幼孢子体,期间可通过涮帘、冲帘等方法防治底栖硅藻污染,底栖硅藻对海带幼体危害不大。而在配子体育苗中,海带配子体小段通过沉降作用附着在苗种帘上,当受到扰动时很容易脱落,因此从配子体泼池到幼孢子体扎根于苗种绳内之前的这段时间,不能够通过常规的涮帘、冲帘等方法防治底栖硅藻污染;期间底栖硅藻会在苗种帘上大量附着繁生,与海带的配子体和幼孢子体竞争营养和生长空间,甚至附着、覆盖到海带幼体上而使其受到危害,导致缺苗脱苗,影响苗种质量。
在海带配子体育苗生产中,配子体泼池大约7 d后开始陆续发育成幼孢子体,9~10 d约90%的配子体发育成孢子体。如果在泼池前将海带配子体于灭菌海水中进行雌雄混合,并进行预发育,则可使配子体泼池后迅速发育成孢子体,从而在与底栖硅藻的竞争中占据优势,抑制底栖硅藻的生长。同时海带配子体提早发育成孢子体后,其能更快扎根苗种绳并附着牢固,继而可以通过冲帘、涮帘的方法防治底栖硅藻。因此海带配子体预发育技术可能可以有效解决底栖硅藻污染问题。
海带配子体预发育与配子体扩增培养尽管都是培育配子体,但配子体扩增培养的目的是让配子体生长、增加配子体生物量、雌雄单独培养。而海带配子体预发育是在配子体扩增培养后,当生物量达到生产需求时,再让配子体进行生殖发育。为进一步提高生殖发育效果,将雌雄配子体混合培养。因此,配子体预发育的条件不能简单套用配子体扩增培养的条件,必须进行试验探索。本文研究海带配子体预发育的适宜光照强度和密度条件及预发育的适宜程度,为海带配子体预发育技术在配子体育苗生产中应用提供技术参考。
1 材料与方法
1.1 海带配子体
海带配子体来自大连海宝渔业有限公司种质库,为经过扩增培养后约300 μm长、呈丝须状的“中宝1号”配子体。
1.2 不同光照强度对海带配子体预发育的影响
用煮沸海水[15]分别配制10.0 mg·L-1 NaNO3-N和2.0 mg·L-1 KH2PO4-P的培养液。取无水鲜海带雌、雄配子体各30.0 g,加入低温培养液,在不高于15℃条件下,用组织破碎机切割至1~6细胞,雌雄混匀后,均分成12份,分别转移到12个500 mL锥形瓶中,往锥形瓶中加培养液至500 mL,使每个锥形瓶内配子体密度为10.0 g·L-1。设置光照强度梯度25.0、37.5、50.0、62.5 μmol·m-2s-1,每个光照条件设置3个500 mL锥形瓶。在光周期12 L∶12 D、温度(13±1)℃条件下充气培养。每日用移液枪从每个锥形瓶中取样至载玻片,在200倍显微镜下分别观察100个雌、雄配子体发育情况。发育情况考察指标为雌配子体发育率、排卵率和雄配子体精囊形成率,若观察到雌配子体小段中有变粗的细胞即视为该配子体小段为发育的配子体。
1.3 不同培育密度对海带配子体预发育的影响
取无水鲜海带雌、雄配子体各37.5 g,按照1.2中的方法切割、混合。按照1∶2∶3∶4的比例分成4份,每份均分到3个500 mL锥形瓶中,并加培养液至500 mL,形成配子体密度为5.0、10.0、15.0、20.0 g·L-1四个组。在光周期12 L∶12 D、室温(13±1)℃、光照强度37.5 μmol·m-2s-1条件下充气培养。配子体发育情况观察同1.2。
1.4 预发育程度对出苗的影响
取无水鲜海带雌、雄配子体各7.5 g,按照1.2中的方法切割、混合后,均分成3份,分别转入500 mL三角烧瓶中,加培养液至500 mL预发育,每瓶中配子体浓度为10.0 g·L-1。预发育条件及观察同1.3。预发育开始前(0 d,即未预发育的海带配子体作为对照组)及预发育后每日从每个三角烧瓶中取样进行出苗测试:用移液枪取1 mL配子体液,转移到9 cm培养皿中,加入15 mL培养液,晃匀,盖上培养皿盖,在光周期12 L∶12 D、室温(13±1)℃、光照强度12.5 μmol·m-2s-1条件下培养,每日观察配子体发育情况和孢子体形成率。
1.5 数据统计与方法
雌配子体发育率=发育变粗的雌配子体小段数/雌配子体小段总数×100%
排卵率=排卵的雌配子体小段数/雌配子体小段总数×100%
精囊形成率= 出精囊的雄配子体小段数/雄配子体小段总数×100%
孢子体形成率=形成孢子体的雌配子体小段数/雌配子体小段总数×100%
用SPSS 23.0 分析试验数据,结果以平均值±标准差表示,并作方差分析,当P<0.05时差异具有统计学意义。
2 结果与分析
2.1 不同光照强度对海带配子体生殖发育的影响
海带配子体预发育过程中,不同光照强度对雌配子体生殖发育率的影响见表1。25.0 μmol·m-2s-1光照组预发育3 d观察到有雌配子体变粗,进行生殖发育,之后变粗雌配子体逐渐增多,17 d所有雌配子体均变粗。37.5 μmol·m-2s-1光照组预发育至2 d有雌配子体变粗,15 d所有雌配子体均变粗。50.0 μmol·m-2s-1和62.5 μmol·m-2s-1光照组预发育1 d即可观察到有雌配子体变粗,之后变粗的雌配子体逐渐增多,14 d两个光照组所有雌配子体均变粗。在25.0~62.5 μmol·m-2s-1光照强度范围内,海带雌配子体均可发育,光照强度越大,雌配子体变粗越早越快,达到100.0%生殖发育的时间越短。但当预发育1 d时,不同光照强度组之间雌配子体变粗比例差异不显著,预发育2~16 d期间,光照强度37.5 μmol·m-2s-1以上组雌配子体变粗比例显著高于光照强度25.0 μmol·m-2s-1组,但光照强度37.5 μmol·m-2s-1以上各组之间差异不显著。
表1 海带雌配子体在不同光照强度下发育率
Tab.1
| 预发育时间/d Advance development time | 发育率/% Development rate | |||
|---|---|---|---|---|
| 25.0 μmol·m-2s-1 | 37.5 μmol·m-2s-1 | 50.0 μmol·m-2s-1 | 62.5 μmol·m-2s-1 | |
| 1 | 0.0±0.0 | 0.0±0.0 | 0.3±0.3 | 0.7±0.3 |
| 2 | 0.0±0.0a | 2.3±0.7ab | 4.0±1.2b | 5.0±1.5b |
| 3 | 1.0±0.6a | 6.7±0.9b | 7.3±2.2b | 9.3±1.4b |
| 4 | 4.7±0.9a | 13.0±1.2b | 15.7±2.9b | 18.0±1.7b |
| 5 | 14.7±2.0a | 26.0±4.0b | 31.0±4.2b | 36.7±3.8b |
| 6 | 37.0±3.2a | 51.3±4.5ab | 54.7±5.7b | 56.3±9.6b |
| 7 | 49.0±2.7a | 63.0±3.8b | 65.7±5.7b | 68.3±2.7b |
| 8 | 58.0±2.0a | 71.0±2.3b | 72.3±6.8b | 74.7±3.2b |
| 9 | 62.0±2.5a | 76.3±1.9b | 77.0±3.1b | 79.0±3.6b |
| 10 | 68.7±3.2a | 84.3±1.8b | 85.0±3.0b | 89.0±2.1b |
| 11 | 76.7±3.7a | 88.0±1.2b | 90.0±1.2b | 91.0±1.7b |
| 12 | 83.7±2.7a | 91.0±1.2b | 94.0±1.0b | 94.7±1.3b |
| 13 | 88.7±3.2a | 95.0±0.6b | 97.0±1.2b | 98.3±0.7b |
| 14 | 91.7±1.8a | 98.0±1.2b | 100.0±0.0b | 100.0±0.0b |
| 15 | 95.0±2.1a | 100.0±0.0b | 100.0±0.0b | 100.0±0.0b |
| 16 | 98.0±0.6a | 100.0±0.0b | 100.0±0.0b | 100.0±0.0b |
| 17 | 100.0±0.0a | 100.0±0.0a | 100.0±0.0a | 100.0±0.0a |
注:同行中标有不同字母表示组间有显著性差异(P<0.05),下表同。
Notes:Data marked with different letters in the same line indicated the significant differences between groups(P<0.05).It was the same in the following tables.
不同光照强度对雌配子体排卵率的影响见表2。25.0 μmol·m-2s-1光照组预发育13 d观察到有雌配子体排卵,之后排卵量逐渐增多;17 d雌配子体排卵比率为(26.0±4.6)%。光照强度37.5 μmol·m-2s-1以上各组均预发育至11 d开始排卵;当预发育17 d时,37.5、50.0、62.5 μmol·m-2s-1光照组排卵率分别为(53.7±2.3)%、(58.3±1.9)%、(59.3±3.8)%。在各组光照条件下预发育,雌配子体均可排卵,光照强度越大,雌配子体排卵越快。排卵率的显著差异主要发生在25.0 μmol·m-2s-1组与其他高光照组之间;37.5、50.0、62.5 μmol·m-2s-1组之间排卵率偶尔有显著差异,但16 d以后无显著差异。
表2 不同光照强度下海带雌配子体排卵率
Tab.2
| 预发育时间/d Advance development time | 排卵率/% Ovulation rate | |||
|---|---|---|---|---|
| 25.0 μmol·m-2s-1 | 37.5 μmol·m-2s-1 | 50.0 μmol·m-2s-1 | 62.5 μmol·m-2s-1 | |
| 11 | 0.0±0.0 | 0.3±0.3 | 1.0±0.7 | 1.3±0.7 |
| 12 | 0.0±0.0a | 2.3±0.7a | 4.3±1.2ab | 5.3±1.2bc |
| 13 | 1.0±0.6a | 6.7±2.4ab | 10.7±2.2b | 12.7±1.8b |
| 14 | 6.0±1.0a | 14.7±2.0b | 27.0±2.3c | 30.0±2.0cd |
| 15 | 12.7±1.5a | 32.3±2.3b | 38.3±2.1bc | 40.0±2.9cd |
| 16 | 18.3±3.5a | 47.7±2.2b | 52.7±2.2b | 54.7±3.8b |
| 17 | 26.0±4.6a | 53.7±2.3b | 58.3±1.9b | 59.3±3.8b |
在海带配子体预发育过程中,不同光照强度对雄配子体精囊形成率的影响见表3。25.0 μmol·m-2s-1光照组预发育4 d观察到有精囊,之后精囊逐渐增多;15 d所有雄配子体小段上均有精囊。37.5 μmol·m-2s-1光照组预发育至4 d观察到有精囊,13 d所有雄配子体均发育出精囊。50.0、62.5 μmol·m-2s-1光照组预均发育3 d观察到精囊,12 d所有雄配子体均发育出精囊。在25.0~62.5 μmol· m-2s-1光照强度范围内,海带雄配子体均可发育出精囊,光照强度越大,雄配子体发育出精囊越早越快,精囊形成率达到100.0%的时间越短。精囊形成率的显著差异主要发生在25.0 μmol·m-2s-1与其他高光照组之间;37.5、50.0、62.5 μmol·m-2s-1预发育组之间精囊形成率偶尔有显著差异,但9 d以后差异均不显著。
表3 不同光照强度下海带雄配子体精囊形成率
Tab.3
| 预发育时间/d Advance development time | 精囊形成率/% Sperm sac formation rate | |||
|---|---|---|---|---|
| 25.0 μmol·m-2s-1 | 37.5 μmol·m-2s-1 | 50.0 μmol·m-2s-1 | 62.5 μmol·m-2s-1 | |
| 3 | 0.0±0.0 | 0.0±0.0 | 1.7±0.7 | 2.0±1.5 |
| 4 | 6.7±0.7a | 10.0±0.6a | 20.3±2.3b | 22.3±3.8b |
| 5 | 27.7±3.8a | 36.0±3.2ab | 47.7±4.3bc | 57.0±6.8c |
| 6 | 40.0±3.0a | 52.0±2.7b | 62.0±2.1c | 73.7±3.5d |
| 7 | 51.0±1.5a | 66.0±2.3b | 78.0±3.2c | 85.0±3.0c |
| 8 | 62.0±2.7a | 75.0±3.1b | 84.7±2.9c | 90.0±1.5c |
| 9 | 71.7±3.2a | 85.0±3.2b | 90.3±1.5b | 92.3±1.9b |
| 10 | 80.7±2.3a | 94.0±1.2b | 97.0±0.6b | 97.7±0.9b |
| 11 | 86.3±3.2a | 97.0±0.6b | 98.0±0.6b | 98.0±1.2b |
| 12 | 91.3±1.8a | 99.3±0.7b | 100.0±0.0b | 100.0±0.0b |
| 13 | 94.7±2.0a | 100.0±0.0b | 100.0±0.0b | 100.0±0.0b |
| 14 | 99.0±1.0 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 |
| 15 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 |
| 16 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 |
| 17 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 | 100.0±0.0 |
在25.0~62.5 μmol·m-2s-1光照强度范围内进行海带雌雄配子体混合预发育,雌配子体均可变粗,进行生殖发育,并排卵,雌配子体均可形成卵囊。光照越强,雌配子体变粗越早越快,排卵越早,雄配子体形成精囊的时间越早,但62.5 μmol·m-2s-1组有个别细胞色素减少、颜色变浅。整体而言,37.5 μmol·m-2s-1光照组与25.0 μmol·m-2s-1光照组的发育差别远大于37.5 μmol·m-2s-1组分别与50 μmol·m-2s-1组、62.5 μmol·m-2s-1组的发育差异。对于海带配子体预发育实际生产操作而言,光照强度越高,操作难度越大,因此光照强度37.5 μmol·m-2s-1是海带配子体预发育的最适宜光照条件。
2.2 不同培育密度对海带配子体预发育的影响
在海带配子体预发育试验中,不同培育密度对雌配子体发育率的影响见表4。 5.0、10.0 g·L-1密度组的海带雌配子体在2 d时均发现有少量变粗发育,之后逐渐增多,5.0 g·L-1密度组在14 d时雌配子体发育率达到100.0%,10.0 g·L-1密度组16 d达到100.0%。15.0 g·L-1密度组在3 d时发现有雌配子体变粗,进行生殖发育,在13 d时39.3%雌配子体变粗发育,而且之后基本维持不变,其余的雌配子体进行丝状体生长,不进行生殖发育。20.0 g·L-1密度组在7 d时有(1.0±0.6)%变粗,之后缓慢增多,15 d时有(14.7±0.3)%雌配子体变粗发育,随后基本维持不变,其余的进行丝状体生长,不进行生殖发育。
表4 不同培育密度下海带雌配子体发育率
Tab.4
| 预发育时间/d Advance development time | 发育率/% Development rate | |||||
|---|---|---|---|---|---|---|
| 5.0 g·L-1 | 10.0 g·L-1 | 15.0 g·L-1 | 20.0 g·L-1 | |||
| 2 | 1.3±0.7a | 2.0±0.6a | 0.0±0.0ab | 0.0±0.0ab | ||
| 3 | 5.0±0.6a | 6.7±1.2a | 2.0±0.6b | 0.0±0.0b | ||
| 4 | 17.0±1.5a | 12.3±1.9b | 5.7±0.9c | 0.0±0.0d | ||
| 5 | 41.0±3.6a | 25.0±4.6b | 11.3±0.3c | 0.0±0.0d | ||
| 6 | 60.0±4.0a | 50.3±3.9b | 15.3±0.3c | 0.0±0.0d | ||
| 7 | 71.7±4.3a | 62.3±4.2a | 22.0±1.7b | 1.0±0.6c | ||
| 8 | 77.0±4.5a | 67.3±1.8b | 23.3±1.3c | 2.3±0.7d | ||
| 9 | 88.7±2.2a | 73.7±3.3b | 25.7±1.8c | 3.7±0.3d | ||
| 10 | 91.7±1.8a | 81.7±2.7b | 30.3±1.5c | 5.3±0.7d | ||
| 11 | 93.0±1.2a | 87.3±1.9a | 37.0±2.7b | 9.0±0.6c | ||
| 12 | 96.0±1.5a | 92.0±2.3a | 38.3±1.8b | 11.3±0.3c | ||
| 13 | 98.0±0.6a | 94.0±1.2b | 39.3±1.8c | 12.0±0.6d | ||
| 14 | 100.0±0.0a | 97.3±0.3a | 40.0±2.3b | 13.3±0.9c | ||
| 15 | 100.0±0.0a | 99.3±0.3a | 39.7±0.9b | 14.7±0.3c | ||
| 16 | 100.0±0.0a | 100.0±0.0a | 39.7±1.2b | 14.7±1.2c | ||
| 17 | 100.0±0.0a | 100.0±0.0a | 40.0±0.6b | 14.3±0.7c | ||
预发育培养的配子体密度越小,雌配子体生殖发育越早越快,预发育效果越好。当配子体密度10.0 g·L-1以下可以完全生殖发育,配子体密度达到15.0 g·L-1及以上时,雌配子体不能完全进行生殖发育。在17 d的预发育试验中,4个密度组之间雌配子体发育率均存在显著差异,但10.0 g·L-1密度组与5.0 g·L-1密度组雌配子体生殖发育率有7 d差异显著,与15.0 g·L-1和20.0 g·L-1密度组雌配子体生殖发育率均有15 d差异显著。
在海带配子体预发育过程中,不同培育密度对雌配子体排卵率的影响见表5。5.0 g·L-1密度组的海带雌配子体在预发育9 d时开始排卵,之后排卵率逐渐增大,17 d时达到95.7%。10.0 g·L-1密度组的海带雌配子体在预发育11 d时开始排卵,之后排卵率逐渐增大,17 d达到51.3%。15.0 g·L-1密度组的海带雌配子体在预发育17 d开始排卵,排卵率为0.3%。20.0 g·L-1密度组的海带雌配子体在本试验的预发育时间内,没有排卵。配子体密度越小,雌配子体发育越好,越容易排卵,当配子体达到15.0 g·L-1及以上时,雌配子体发育不理想,不容易排卵。雌配子体开始排卵后,5.0 g·L-1密度组排卵率均显著高于其余各组。10.0 g·L-1密度组13 d之后的排卵率均显著高于15.0 g·L-1和20.0 g·L-1组。
表5 不同培育密度下海带雌配子体排卵率
Tab.5
| 预发育时间/d Advance development time | 排卵率/% Ovulation rate | |||
|---|---|---|---|---|
| 5.0 g·L-1 | 10.0 g·L-1 | 15.0 g·L-1 | 20.0 g·L-1 | |
| 9 | 0.7±0.3a | 0.0±0.0b | 0.0±0.0b | 0.0±0.0b |
| 10 | 6.0±1.2a | 0.0±0.0b | 0.0±0.0b | 0.0±0.0b |
| 11 | 15.3±0.3a | 0.3±0.3b | 0.0±0.0b | 0.0±0.0b |
| 12 | 19.3±3.0a | 2.0±0.6b | 0.0±0.0b | 0.0±0.0b |
| 13 | 38.7±1.8a | 6.0±2.1b | 0.0±0.0c | 0.0±0.0c |
| 14 | 54.0±2.7a | 13.7±1.2b | 0.0±0.0c | 0.0±0.0c |
| 15 | 69.0±2.7a | 30.0±2.1b | 0.0±0.0c | 0.0±0.0c |
| 16 | 84.3±2.3a | 47.0±2.5b | 0.0±0.0c | 0.0±0.0c |
| 17 | 95.7±0.9a | 51.3±2.4b | 0.3±0.3c | 0.0±0.0c |
在海带配子体预发育过程中,不同培育密度对雄配子体精囊形成率的影响见表6。5.0 g·L-1密度组的海带雄配子体在预发育3 d时开始出现精囊,之后精囊形成率迅速提高,12 d达到100.0%。10.0 g·L-1密度组在预发育4 d时开始出现精囊,之后精囊形成率迅速增长,13 d达到100.0%。15.0 g·L-1密度组在预发育5 d时开始出现精囊,之后精囊形成率呈较慢增长,17 d达到86.7%,少量雄配子体进行丝状体生长,配子体加长,未进入生殖发育过程。20.0 g·L-1密度组在预发育8 d时才开始出现精囊,之后精囊缓慢增多,17 d达到27.7%,大部分雄配子体进行丝状体生长。配子体密度越小,雄配子体发育越好,越容易形成精囊,当配子体达到15.0 g·L-1及以上时,部分雄配子体进行丝状体生长,不形成精囊。
表6 不同培育密度下海带雄配子体精囊形成率
Tab.6
| 预发育时间/d Advance development time | 精囊形成率/% Sperm sac formation rate | |||
|---|---|---|---|---|
| 5.0 g·L-1 | 10.0 g·L-1 | 15.0 g·L-1 | 20.0 g·L-1 | |
| 3 | 6.3±0.9a | 0.0±0.0b | 0.0±0.0b | 0.0±0.0b |
| 4 | 27.0±2.7a | 10.0±1.2b | 0.0±0.0c | 0.0±0.0c |
| 5 | 46.0±3.6a | 34.7±2.6b | 2.0±0.6c | 0.0±0.0c |
| 6 | 54.7±3.2a | 50.0±2.5a | 17.3±1.2b | 0.0±0.0c |
| 7 | 67.3±2.9a | 65.0±2.7a | 22.3±1.8b | 0.0±0.0c |
| 8 | 85.7±1.9a | 76.7±3.2b | 31.7±3.4c | 1.3±0.9d |
| 9 | 92.3±1.3a | 86.0±2.1a | 43.3±3.4b | 3.3±0.9c |
| 10 | 96.7±0.9a | 94.3±0.3a | 50.0±2.9b | 6.3±0.9c |
| 11 | 99.3±0.7a | 98.0±0.6a | 62.0±1.7b | 11.7±1.8c |
| 12 | 100.0±0.0a | 99.3±0.7a | 64.3±1.2b | 15.7±1.8c |
| 13 | 100.0±0.0a | 100.0±0.0a | 73.0±2.7b | 18.0±2.0c |
| 14 | 100.0±0.0a | 100.0±0.0a | 74.7±2.3b | 22.0±1.7c |
| 15 | 100.0±0.0a | 100.0±0.0a | 78.0±1.0b | 24.3±1.2c |
| 16 | 100.0±0.0a | 100.0±0.0a | 80.0±1.5b | 26.0±1.2c |
| 17 | 100.0±0.0a | 100.0±0.0a | 86.7±1.2b | 27.7±1.5c |
在17 d的预发育过程中,10.0 g·L-1密度组与5.0 g·L-1密度组的精囊形成率只有4 d差异显著,预发育9 d后,精囊形成率差异均不显著;而10.0 g·L-1密度组在出现精囊后,其精囊形成率与15.0 g·L-1密度组和20.0 g·L-1密度组均存在显著差异。
在培育密度达到15.0 g·L-1及以上条件下预发育,海带雌雄配子体容易进入丝状体生长状态,但发育效果不理想。在5.0、10.0 g·L-1密度下预发育,雌雄配子体均能进行正常生殖发育,但10.0 g·L-1组发育速度略慢于5.0 g·L-1。预发育时,配子体密度越大,单位体积培养液可预发育的配子体越多,生产效率越高,因此10.0 g·L-1密度为海带配子体预发育的适宜密度。
2.3 预发育程度对出苗的影响
在光照强度37.5 μmol· m-2s-1、光周期12 L∶12 D、室温(13±1)℃、密度10.0 g·L-1条件下进行海带配子体预发育,于不同预发育时间取样进行出苗测试(表7)。预发育0 d,其出苗率达到90.0%以上的培育时间为9 d。随着预发育时间增加,出苗率达到90.0%以上的时间越短,预发育10~14 d各组的出苗率达到90%以上所需的培育时间最短,只要5 d。预发育12 d的配子体,培育5 d的出苗率最高,达到96.7%(图1)。预发育时间超过16 d后,出苗率达到90.0%以上的时间逐渐增加;预发育21 d组的出苗率最大只有90.0%,而且需要8 d培育时间。预发育22 d以后,最大出苗率均达不到90.0%。后续试验还发现,预发育26 d以后,雌、雄配子体颜色均变浅,最大出苗率只有(18.5±7.3)%,且苗体颜色浅、部分透明。预发育28 d之后,雌、雄配子体部分发白,变成空腔死亡,只发育出少量幼苗,且陆续发白死亡。因此海带配子体预发育时间不宜过长,预发育的最适宜时间为11~12 d,比未预发育配子体提早4 d达到90.0%以上出苗率。
表7 预发育时间对出苗效果的影响
Tab.7
| 预发育时间/d Advance development time | 出苗培育时间/d Cultivation time for emergence | 出苗率/% Emergence rate |
|---|---|---|
| 0 | 9 | 90.7±1.2 |
| 1 | 9 | 92.0±1.2 |
| 2 | 9 | 95.3±1.8 |
| 3 | 8 | 90.3±0.3 |
| 4 | 8 | 93.0±1.2 |
| 5 | 8 | 95.0±1.2 |
| 6 | 7 | 91.7±0.7 |
| 7 | 7 | 95.3±0.7 |
| 8 | 6 | 90.3±0.7 |
| 9 | 6 | 96.3±0.9 |
| 10 | 5 | 92.3±0.9 |
| 11 | 5 | 96.3±1.2 |
| 12 | 5 | 96.7±0.9 |
| 13 | 5 | 95.3±0.7 |
| 14 | 5 | 94.3±0.3 |
| 15 | 6 | 93.0±0.6 |
| 16 | 6 | 92.7±0.3 |
| 17 | 7 | 92.0±0.6 |
| 18 | 7 | 90.0±0.6 |
| 19 | 7 | 91.0±0.6 |
| 20 | 7 | 90.0±0.6 |
| 21 | 8 | 90.0±0.6 |
| 22 | 8 | 89.7±0.3 |
| 23 | 8 | 85.3±0.3 |
图1
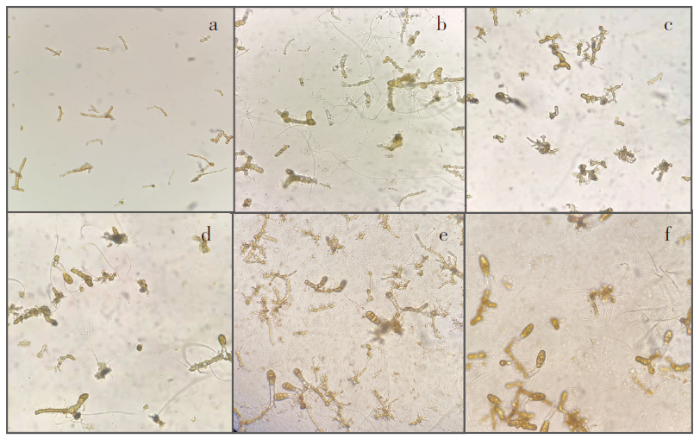
图1
海带配子体预发育及其出苗过程
注:a.切割后预发育前的配子体;b.预发育6 d配子体;c.预发育12 d配子体;d.出苗培育3 d;e.出苗培育4 d;f.出苗培育5 d。
Fig.1
Gametophyte advance development and emergence of Saccharina japonica
Notes:a.Fragment of gametophyte before advance development;b.Gametophyte of advance developed for 6 days;c.Gametophyte of advance developed for 12 days;d.Seedling cultivation for 3 days;e.Seedling cultivation for 4 days;f.Seedling cultivation for 5 days.
3 讨论
3.1 光照强度对海带配子体发育的影响
陈书秀等[4]研究了笼目海带(Kjellmaniella crassifolia Miyabe)配子体发育与光照强度的关系,认为光照强度对笼目海带配子体的发育有显著的影响,笼目海带配子体发育的最适光照强度为20~40 μmol·m-2s-1 。Lüning K[16]研究表明,当海带配子体在充足光照下会进入生殖发育,在低光下则维持营养生长状态。Lee J A[17]研究表明,当光照强度为80 μmol·m- 2 s-1 时,皱海带(Laminaria religiosa) 配子体的繁殖率达到最大。杨迎霞等[5]研究结果表明,在56.3 μmol·m- 2 s-1 光照强度下,充气悬浮培养的海带配子体克隆可较快生长发育,但尚书[18]认为,在56.3 μmol·m- 2 s-1 光照强度下低密度(0.1 g·L-1)培养海带配子体,容易造成细胞色素淡化。本研究结果表明,在光照强度25.0 ~ 62.5 μmol·m-2s-1范围内,培育密度为10.0 g·L-1的海带配子体,光照越强,发育越快,37.5 μmol·m-2s-1光照条件预发育效果显著优于25.0 μmol·m-2s-1,与50.0~62.5 μmol·m-2s-1光照条件的培育效果差异不显著。62.5 μmol·m-2s-1光照强度会使个别配子体颜色变淡。育苗生产中扩增培养配子体时,一般利用多层培养架,每层培养架摆放2排培养瓶,上方设置2~3排日光灯管作为光源。预留出充气管的操作空间后,锥形瓶液面位置的光照强度可达到40 μmol·m-2s-1左右。若需要较低的光照强度,可以遮蔽部分日光灯管。但若要提高光照强度,则需要改换更大功率的日光灯管,同时日光灯管释放的热量也会增多,不利于控温操作。因此考虑到实际生产中可行性和经济适用性,37.5 μmol·m-2s-1可作为海带配子体预发育的理想光照强度。
3.2 培育密度对海带配子体发育的影响
本研究发现,配子体密度越小,雌配子体发育率、排卵率、雄配子体精囊形成率越高。当培育密度达到15.0 g·L-1及以上时,配子体不易进行生殖发育,而容易进行丝状体生长,达不到预发育的效果。高密度培养影响海带配子体预发育的原因可能有2个方面:一是高密度培养限制了配子体受光时间和受光量。本研究中,海带配子体是在锥形瓶中充气翻腾培养,培育密度大,会使单位配子体光照强度不足,受光时间变短,影响甚至抑制生殖发育。而该受光时间和受光量可能正是海带配子体进行丝状体生长的适宜条件,因此配子体进入生长过程,而非生殖发育过程。海带配子体生殖发育和丝状体生长的条件应该存在明显差异,但机理有待进一步的试验研究。二是在高密度培养条件下,培养液中海带配子体生殖发育必需的营养成分,比如铁离子[19-20]等可能不足,限制了预发育效果。
3.3 预发育时间对海带配子体出苗的影响
本研究中,在37.5 μmol·m-2s-1、光周期12 L∶12 D、室温(13±1)℃、密度10.0 g·L-1条件下预发育12 d的海带配子体,出苗培育5 d,出苗率达到(96.7±0.9)%,比未预发育的配子体提早4 d达到90.0%以上的出苗率。但预发育时间太长,预发育效果会降低甚至失败。过度预发育导致预发育失败的原因可能是雌雄配子体发育到一定程度后,在相对较小的空间内继续发育致使环境条件不能满足需求,同时其代谢物可能也会对配子体造成伤害,最终导致雌雄配子体和后续形成的幼苗死亡。因此进行海带配子体预发育需要严格控制预发育的程度,以保证配子体和苗种的质量。
3.4 配子体预发育状态指标的选择
本研究以雌配子体发育率、排卵率、精囊形成率为考察指标,观察海带配子体预发育的状态和效果。其中雌配子体排卵率和雄配子体精囊形成率是常用的观察指标,用这两个指标表述配子体发育的结果。而配子体的发育过程用雌配子体发育率来考察。配子体开始发育以雌配子体变粗作为标志,若观察到雌配子体小段中有变粗的细胞即视为该配子体小段为发育的配子体。本研究中以配子体发育率考察配子体的发育过程,属首次使用。当雌配子体变粗时,配子体形态发生变化,可以通过显微镜分辨出来。不同配子体在发育之前的粗细不完全一致,变粗之后的粗细也不都是一个规格,因此难以用量化的粗细变化作为判断标准,只能做定性考察。 雄配子体发育过程中,细胞也有一定程度的变化,但不如雌配子体变粗更易于观察。更科学严谨的配子体发育过程的观察指标有待在今后的研究中进一步开发。
4 结论
海带配子体预发育的适宜光照强度为37.5 μmol·m-2s-1、适宜密度为10.0 g·L-1、适宜预发育时间为11~12 d。
参考文献
光照强度和光照周期对笼目海带配子体发育的响
[J].
通过设置不同的光照强度和光照周期,在实验室条件下检测了笼目海带(Kjellmaniella crassifolia Miyabe)配子体的发育情况。结果显示:光照强度及光照周期均对笼目海带配子体的发育率有显著的影响(P<0.05)。结果表明:1)在光照强度条件不同、其他培养条件相同的情况下,经过4周的发育,5~80 μmol·m-2s-1条件下大部分配子体进入了生殖生长状态,发育率为60%~80%,其中20~40 μmol·m-2s-1光照强度条件下生殖生长明显快于其他组。2)在光周期不同、其他培养条件相同的情况下,培养第12~25天时,20L∶4D条件下发育率为6.68%~69.87%,发育率与16L∶8D实验组差异不显著,但显著大于其余组。培养第32 天,光照时间最长的处理组即20L∶4D的发育率(74.54%)最高,且孢子体细胞列数最多。本研究为笼目海带配子体规模化扩增及育苗提供了数据参考。
On the temperature in relation to the development of the gamctophyte of Saccharina religiosa
[J].
An investigation by cultural methods of some of the factors influencing the development of the gametophytes and early stages of the sporophytes of Saccharina digitata,S.digitata and S.saccharina
[J].
Untersuchungen über Parthenogenesis,Geschlechtsbestimmung und Bastardierungsvermögen bei Laminarien
[J].
Interactions of daylength,temperature and nutrients affect thresholds for life stage transitions in the kelp Saccharina digitata(Phaeophyce)
[J].
Critical levels of light and temperature regulating the gametogenesis of three Laminaria species(Phaeophyceae)
[J].
Gametogenesis and early sporophyte development of Laminaria religiosa Miyabe in the east coast of Korea
[J].

{kind=link}
{kind=link}